
1.Что такое строматолит и чем определяется его форма?- такой вопрос одним из первых поставил сотрудник ПИНа АН СССР А.Г.Вологдин и сам, отвечая на него, написал следующее: «мы выделяем форму с куполовидным характером наслоений…, но чем объяснить ту или иную правильную геометрическую форму всего образования мы не знали, да по правде говоря, этим и не занимались»[3, с.131]. Аналогичное высказывание сотрудника группы ученых ГИНа АН СССР В.А.Комара, подтверждает, что «использование морфологических особенностей строматолитовых построек базируется на не доказанном, но обязательном положении о прямой связи между формой строматолитовой постройки и составом водорослей-строматолитообразователей» [14, с.53]. Эти высказывания основывается на представлениях В.П.Маслова 50-х годов, который сделал предположение, что форма строматолитов как, водорослевых образований, зависит от состава водорослей – «хозяев» постройки, как он их назвал, а состав самих «хозяев» определяется фациальными условиями их произрастания в древнем бассейне [16,c.24]. «Откровенные» высказывания известных исследователей строматолитов убеждает нас, что вся концепция формы строматолитов действительно не имела под собой твердой научной основы.
Однако, отсутствие научной теории, объясняющей происхождение формы строматолитов, не помешало тогда И.Н.Крылову, В.А.Комару, М.Е.Раабен и др. их последователям «взять предположение В.П.Маслова за основу» и рассматривать строматолиты как «видоспецифические» колониальные водорослевые образования и применять их для стратиграфии докембрия. Такое чисто эмпирическое уравнивание строматолитов с обычными видами организмов затем рассматривается как оправдание применения к ним линнеевской бинарной номенклатуры, которая к колониальным органическим постройкам типа биогермов еще никем не применялась. При этом приводились ссылки на ботанический кодекс, якобы допускающий подобную процедуру [15]. Упорство палеонтологов в этом вопросе подозрительно. Другой вариант оправдания бинарных латинских названий, базируется на идее Ф.Я.Власова [5], толковать строматолиты как особый тип организмов вроде лишайников с выделением у них классов, семейств и т.д. Эту идею, как новую, недавно опять выдвинул К.Н.Конюшков, не смотря на то, что она была отвергнута еще в 60-е годы. По-видимому, все дело именно в линнееевской номенклатуре, предусматривающей авторство для каждого вида, рода и любого таксона (см ниже стр.9).
Для случая симбиотических цианобактериальных построек, такие попытки применения ботанического кодекса кажутся не законными. Посудите сами, что предусматриваемое кодексом отнесение сходных по жилкованию листочков, семян или спор неизвестных растений или коры кноррий и лепидодендронов в одну условную группу, не является тождественной операцией с отнесением похожих строматолитовых столбиков в один формальный род. В первом случае мы имеем дело с частями организмов, которые при нахождении целого экземпляра растения будут точно установлены. В случае строматолитов такой надежды быть не может. В первом случае имеем дело с частями растения, имеющих явно конституционную, т.е. генетически обусловленную морфологию объектов, а в случае строматолита имеем только геометрически (морфографически) сходные (похожие) формы колониальных наслоений, а вовсе не частей особи организма. Строматолит это совсем другая надорганизменная (колониальная или ценотическая целостность), с другим — негенетическим способом морфогенеза, на который правила ботанического кодекса не распространяются.
Последующее полвека изучения строматолитов совершенно ясно доказало, что седиментационные процессы на мелководьях древних бассейнов и гидродинамика среды являются главными факторами, определяющими форму строматолитовых построек и поэтому форма их не видоспецифична, а скорее экоспецифична [16,25,29].
Расцвет ложной идеи «видоспецифичности» форм строматолитов и игнорирования явной зависимости их формы от среды обитания является главной причиной неудачи при построения их естественной классификации и сомнительного применения в стратиграфии докембрия. Это ведь только советские и российские ученые верят в строматолитовые комплексы якобы характеризующие отдельные стратиграфические подразделения докембрия. Как показал Геологический конгресс в Киото, многие зарубежные геологи, например конвинер Палеонтологического симпозиума Адольф Цейлахер из Тюбингена, очень даже сомневаются в строматолитовой стратиграфии докембрия и считают, что в данном случае желаемое выдается за действительное [40].
Дело в том, что конституционный, видоспецифический (генный) или биологически — зависимый морфогенез форм индивидуальных организмов (особей) тоже в значительной степени зависит от экологических факторов и формирует адаптированные к среде формы организмов, но их морфогенез или точнее сказать его норма, закреплен в генетической конституции организма[1], о чем разъясняется в кодексе. Однако существует еще одна проблема- это проблема морфогенеза форм сложных колониальных организмов и проблема морфопоэза их ценотических образований, какими являются такие сложные образования как строматолиты, (или как их удачно обозвала А.И.Антошкина — цементолиты) у которых генетический, конституционный морфогенез не играет роли.
Отсутствие в биологии теории морфогенеза таких сложных форм надорганизменных образований имело плачевные последствия для палеонтологии строматолитов. Последние стали изучаться более ста лет назад именно потому, что как органические образования, они потенциально могли иметь стратиграфическое значение и оказать помощь в расшифровке геологической истории докембрийского этапа развития Земли, лишенного обычных органических остатков (преимущественно многоклеточных эукариотических организмов), свойственных фанерозою. Принимая трактовку форм строматолитов как форм построек колониальных организмов следует вникнуть в проблему морфогенеза колоний. А этого никто, кроме автора данного текста, делать не захотел, и проблема с тех пор повисла в воздухе, поскольку в палеонтологии нет никакой теории формы строматолитов. Как и 50 лет назад [23,24], я еще раз пытаюсь показать, что вопреки мнению Фентонов, А.Г.Вологдина, К.Б.Корде и прочих, решение проблемы существует, но не на том пути, по которому пошли В.П.Маслов, И.Н.Крылов и их последователи, а на пути освоения учения о жизненных формах.
- Изучение истории проблемы морфогенеза биологических форм, показывает, что еще в 1919 году Твенхофел предложил для обозначения элементарных водорослевых слоев-колоний термин — ценоплаз, а в 1927 г. Юлиус Пиа [37] предложил рассматривать формы строматолитов как формы роста водорослевых колоний (wichtigchte Wuschtypen). Аналогичный термин — Life-forms (жизненные формы) применили в своей статье 1931 года и Фентоны.
Исключением из патовой ситуации с биоморфологией являются одиночные попытки построения общей теории происхождения биологической формы в проморфологических исследованиях В.Н.Беклемишева [2], механо-морфологических исследованиях А.А.Еленкина [10], биополевых А.Г.Гурвича [8], поставивших проблему в общем плане. Связь биологического морфогенеза с генетикой эукариот показали работы Конрада Уоддингтона [32]. Морфогенезом растений занимался Эдмунд Синнот [21]. Некоторые эколого- морфологические наблюдения за ростом колониальных кораллов в Красном море, видимо одним из первых в 1913 г, произвел Н.Н.Яковлева [34], затем Д.В.Наумов, Кин-сун и Хуан Мин-сянь в Желтом море [17], Юджин Шинн у побережья Флориды [39]. На ископаемом материале (на колониальных кораллах) такие исследования осуществили Б.В.Преображенский [18] и У.Оливер [38]. Все указанные исследователи однозначно показали сильное влияние формирующей роли среды на форму не только самих организмов, но особенно на форму их колоний, биогермов и рифовых построек или «цементолитов», аналогов строматолитов.
С начала 80-х годов к изучению биохимии и физиологии сложных цианобактериальных сообществ подключилась большая группа микробиологов, позволившая значительно продвинуть понимание сложных процессов биохимических и трофических связей, существующих в надорганизменных полиморфных сообществах вообще и в строматолитовых в частности [12]. Их исследования неожиданно проливают новый свет и на морфогенетическую проблему сложных цианобактериальных образований. Они убедительно доказывают существование различных степеней интеграции в биологических системах колониального и ценотического уровней организации. Единственной теорией способной описать морфогенез биосистем различного уровня интеграции от особей до колоний и ценотических построек является эколого-морфологическая теория жизненных форм [25,27].
Прижизненная форма цианобактериальной колонии и ископаемая форма строматолитового столбика [16, c.46], состоящего из стопки элементарных слоев в «классическом подходе» [15] не различаются. Видимая на выветрелой поверхности породы или восстановленная графическим препарированием ископаемая форма строматолитов имеет метахронную структуру, но она не тождественна форме организма или колонии. На рисунках И.Н. Крылова, обратите внимание, не показывается даже куполообразная поверхность элементарного слоя, самый главный диагностический признак живой колонии, все внимание отдано форме столбиков, т.е. ископаемой форме. Возникает вопрос – понимал ли он то, что делал, увлекшись графическим препарированием и понимают ли все его последователи как следует описывать и классифицировать строматолиты?
Толкование форм строматолитов как форм роста водорослевых колоний, предложенное Юлиусом Пиа [37], касалось именно формы элементарных прижизненных слоев. Двоякое понимание формы колоний запутывает дело. Более того, термины «life-forms» и «wichtichgste Wuchstypen», как видно из практики их применения в нашей стране, ни о чем не говорили и были терминами свободного пользования. Это не давало возможности выйти из теоретического тупика, т.к. в биологической систематике не существовало и не существует классификаций колониальных организмов и отсутствует толкование термина «форма роста» в морфологическом смысле.
Учение о жизненных формах, изобретенное Александром фон Гумбольдтом [6,20,23] оказалось такой подходящей теоретической базой, приемлемой для описания водорослевых колоний и для адекватного толкования формы строматолитов. Так, например, с помощью экологической морфологии очень просто решался парадокс с формами кембрийских строматолитов хребта Каратау, с которым столкнулся И.Н.Крылов [15, c.73-86]. Прекрасная сохранность упомянутых строматолитов показала, что формы столбиков, взятых из различных частей одного биогерма: из основания (базо-формы), из кровли (крышка-формы), из центра (центро-формы) (здесь в скобках термины И.Н.Крылова) или из его краевых частей, различаются между собой больше, чем различные «роды» или формальные группы строматолитов. Так, формы, находящиеся в основании биогерма, оказались эквивалентными пластовым формам Stratifera или пластово–столбчатым формам типа Columnaefacta или Kussiella, краевые столбики – соответствовали диагнозу Tungussia, а формы столбиков из центральных частей биогерма в точности подходили под диагноз Gymnosolen, хотя все они были созданы заведомо одним и тем же ценозом или сообществом цианобактерий, построивших описываемый биогерм. Это наблюдение убедительно показывает, что формальный диагноз, основанный на «морфографическом» (или иконографическом) принципе при описании и выделении «родов и видов» строматолитов не дает реальных естественных единиц. Тем более, как можно к водорослево-бактериальному биоценозу применять понятие рода или вида в линнеевском понимании и давать им латинские названия?! Это явная псевдонаука в чистом виде.
Необходима другая система представлений на морфологию строматолитов. Согласно предложенной здесь эколого-морфологической теории форм роста или теории жизненных форм, все упомянутые различные формы столбиков являются просто экоморфно- подобными постройками или формами роста- экологическими разновидностями одной жизненной формы или постройки бактериально-водорослевого ценоза. Экоморфа симбиотической колонии или ценотического желвака, в отличие от ЖФ, есть результат адаптации симбиотического комплекса организмов к какому-то одному фактору среды, например, краевому положению в биогерме, в этом случае формируются тунгуссоидные формы, а в центре биогерма образуются формы типа гимносоленид, в подошве биогерма – слоистые стратиферы и т.д.
Опасность и нежелательность формального морфографического (геометрического) подхода к строматолитам состоит в том, что любое незначительное изменение вида столбиков, описывается как новый вид или род без всяких оснований, что ведет к засорению не только систематики, но и геологической литературы лженаучной красивой латинизированной мишурой. По данным М.Е. Раабен к началу 2000 года уже было описано около 1500 таксонов различного ранга, из них более 90% родовых и более 1000 видовых единиц, странно, что такая «плодовитость» строматолитчиков не вызвала ни у кого подозрения [19]. Думается, что наш классик В.О. Ковалевский в письме из Берлина своему брату А.О. Ковалевскому (весна 1870 г.) именно относительно такой палеонтологии писал, что «кроме пошлого описания пуплышек и ребрышек в ней ничего нет, так что стыдно читать» [9, с.191-212]. Известный исследователь трилобитов Ричард Форти раскрывает нам психологическую подоплеку стремления палеонтологов, и строматолитчиков в том числе, применять линнеевскую бинарную номенклатуру к ископаемым формам жизни. Он объясняет это тем, что какой-то совсем безвестный А.Б.Иксов, нашедший новый вид трилобита, благодаря правилу линнеевской систематики сразу попадает в один ряд с великими естествоиспытателями как Карл Линней, Иоахим Барранд или Чарльз Дарвин, чьи имена навеки вошли в анналы науки, кроме всего прочего, еще в качестве авторов когда-то впервые описанных ими видов[41]. Подсчеты М.Е.Раабен показывают сколько появилось таких желающих навечно вписать свои имена буквально ни на чем, описывая разные ветки дерева.
- Исследование зависимости форм строматолитов от фаций, начатые К.Л и М.А.Фентонами [42] и В.П.Масловым [16] были продолжены мной на материале кунгурских строматолитов Актюбинского Приуралья [7,24]. Прослеживая изменение их форм по профилю вдоль древней береговой линии и в прибрежных фациях актастинской пачки известняков В.П.Пнева, я обнаружил определенную зависимость между геологическими фациями (условиями среды осадконакопления), с одной стороны, и характером форм строматолитов- с другой (рис 1). Оказалось, что один и тот же цианобактериальный комплекс формирует разные формы строматолитов в различных фациях, а различные комплексы цианобактерий в сходных условиях образуют сходные формы строматолитов. Морфологическое сходство форм, создаваемых различными сообществами цианофицей и бактерий, определяется законом экологической конвергенции в одинаковых гидродинамических и гидрохимических режимах водоема. Следовало выяснить какие факторы среды и как определяют форму строматолитов, но на рифейском материале такие исследования, как утверждает И.Н.Крылов, не проводились. Строматолиты при этом должны рассматриваться как морфологически целостные симбиотические колонии (надорганизменные образования колониального уровня) и как цианобактериальные синцианозы (термин А.А.Еленкина) или «маты» (по терминологии американских альгологов и отечественных микробиологов), т.е. как водорослево-бактериальные надорганизменные образования ценотического уровня организации живой материи [23,25]. Априорно, из общих соображений, следовало ожидать, что строматолиты колониального уровня и строматолиты ценотического уровня должны несколько по- разному взаимодействовать со средой и подчиняться различным факторам морфогенеза.
Среди основных факторов экологического морфогенеза строматолитов необходимо рассмотреть следующие: фактор абсолютного размера форм, корреляции роста, полярность форм и проблему целостности строматолита как полифитного образования с системой зависимых физиологических и биохимически-сопряженных реакций карбонатоосаждения. Без решения проблемы целостности строматолита все разговоры об их форме не имели под собой научной основы. Именно поэтому В.П.Маслов интуитивно, а И.Н.Крылов сознательно, вводили термин и понятие о «водорослях-хозяевах», что, как теперь выясняется, было ошибкой. Понятие жизненной формы исправляет эту ошибку и адекватно отражает суть форм строматолитов.
Фактор абсолютного размера определяет пределы роста выпуклых колониальных форм через критическую величину отношения питающей поверхности к объему живого вещества. У строматолитов он определяет начало деления (распадения) одной колонии на две или несколько дочерних (например, пассивное распадение одного столбика у Kussiella на два дочерних). С увеличением размера колонии ее поверхность — S растет пропорционально квадрату линейных размеров или радиуса — R, а объем пропорционально кубу того же параметра. Иными словами, кривизна поверхности колонии при ее росте уменьшается как 1/R. По закону Л.Берталанфи для биологических систем организменного уровня интеграции это определяет критическую величину отношения интенсивности процессов ассимиляции и диссимиляции, при которой питающая поверхность уже не в состоянии обеспечить питанием растущую массу живого вещества организма, а в нашем случае — колонии. Начавшая голодать колония распадается на две дочерних, у которых параметр 1/R снова становится достаточно высок и рост колонии может продолжаться. Этот универсальный закон роста Герберт Спенсер впервые применил для объяснения предела роста клеток, а Джулиан Гексли для объяснения аллотропного характера роста органов и частей организмов. Что мешает нам использовать его для объяснения роста и деления колоний или ископаемых строматолитов? Ничего! Почему же никто из строматолитчиков, имея такой обильный каменный материал, им не воспользовались?
Далее, второй закон геометрического отбора, открытый нашим известным минералогом Д.П.Григорьевым, можно использовать для описания роста колоний хроококковых цианофицей [25], также может быть отнесен к таким универсальным геометрическим законам роста и факторам отбора растущих колоний в борьбе за пространство (рис.2).
Корреляции роста хорошо просматриваются на утолщениях слойков на вершинах строматолитовых форм и утоньшении их на боковых поверхностях, являются прямым следствием закона аллотропного роста Джулиана Гексли. Рост каждого биогерма контролируется таким образом, что составляющие его строматолитовые столбики приобретают разную форму. Открытие наличия внутри биогерма разных форм столбиков, относимых И.Н.Крыловым к разным «родам и видам» строматолитов сразу показало несовершенство его морфографической практики описания и выделения новых форм (более подробно об этом — выше).
Полярность форм прослеживается у всех колоний и построек как неравнозначность концов оси и предпочтение роста в одном направлении, определяемом градиентом поля силы тяжести, степени освещенности или направлением течений. Общенаучный или общебиологический принцип Пьера Кюри позволяет понять часто наблюдаемую псевдогексагональность форм у конофитонов, а способность цианофицей к фототаксису, согласно А.Г.Вологдину, объясняет их коническую форму с присущей им зоной смятия в апикальной части и их наклонный рост в плоскости древнего меридиана [4].
Влияние факторов гидродинамики и осадконакопления на форму строматолитов настолько очевидно, что отмечалось И.Н.Крыловым [15] и описано мной при рассмотрении рифельных (похожих на знаки ряби) форм строматолитов (рис.3) и при описании реотропного роста столбиков, образуемых нитчатыми цианофицеями Tabantalia (рис.2)[27] в строматолитах Tubistromia и Alterogophyton (рис.4).
Физиологическая целостность и сопряженность катаболических и анаболических процессов в водорослевых колониях описана многими ботаниками и микробиологами. Результаты изучения особенности карбонатоосаждения бактериально-циановыми, сообществами начиная с Г.А.Надсона и Б.В.Перфильева вплоть до работ Г.А.Заварзина теперь общеизвестны, хотя и не всегда осознаются как существенное доказательство целостности строматолита или важнейшего интеграционного фактора морфогенеза [25,28].
Седиментологический фактор. Осаждение дисперсных частиц, выпадающих из водной среды на поверхность цианобактериальных пленок, также является мощным экологическим фактором морфогенеза, определяющего рельеф строматолитовых тел. Самоочищение поверхности живых пленок осуществляется путем мерцательных движений нитей и скольжения клеток цианей, а также с помощью волнообразных движений слизистых пленок самих бактерий, подобно тому, как это делают клетки эпителиев дыхательных органов животных. Поверхность живого сообщества или колонии должна непрерывно освобождаться от инородных частиц для осуществления фотосинтеза и обменных процессов со средой, и поэтому вся поверхность цианобактериального сообщества или мата делает это путем сбрасывания чужеродного осадка в промежутки между спонтанно возникающими куполами колоний — (ценоплазами Твенхофела) или в мозговидные извилины или фоссулы на его поверхности. Именно таким путем формируются колонки строматолитов типа Gymnosolen Steinman и мозговидные колонии типа Fosella Maslov или похожие на волновую рябь постройки типа Pitella Komar и прочие строматолитовые поверхности, во впадинах которых скапливается инородный осадочный материал. Даже продукты деструкции собственного биогенного материала — слизистого гликокаликса, регулярно отмирающего после каждого сезона вегетации и сползающие с куполов пермских Gantifera granulose влияют на формообразование строматолитовых тел, порождая «непонятные» и никем, кроме [30] не объясняемые до сих пор столбики (рис.5). Справедливости ради упомянем, что впервые такую самоочищающую способность бактериальных колоний открыли сотрудники знаменитой экспедиции «Челленджера» еще в 1870-х годах, обратившие внимание на «непотопляемость» железо-марганцевых конкреций в осадке дна океана [27,28].
- Поэтому вслед за В.П.Масловым, я считаю необходимым, выделять два уровня организации строматолитовых образований: ценотический, так называемые литостромы, образуемые природными ценозами цианофицей и колониальный, куда следует относить целостные симбиотические образования надорганизменного уровня, называемых колониями, имеющие собственные (идиоморфные) формы типа Collenia, Fosella, Conophyton, Osagia и др. [16]. Отдельно должны описываться формы облекания- (перифитоновые строматолиты, обнаруженные мной в Актюбинском Приуралье), образующиеся разными перифитоновыми сообществами цианобактерий в особых экологических условиях и формирующие на различных субстратах над дном водоема различные формы строматолитов [22,28].
На затопленных в водоеме ветках и стволах деревьев возникают формы типа Tubistromia и Alterogophyton, повторяющие сложную корявую форму веток и стволов и несущие отпечатки сучков и коры либо лепидодендронов, либо каламитов [22,26]. Как отдельные пластовые формы описываются рифельные формы Rifelina, Metarifelina (рис.4) и доскообразные постройки- Pitella и Platella, похожие на знаки ряби обычных мелководий. Формы Pitella, по моим наблюдениям, возникают путем разделения сплошной цианобактериальной пленки, покрывающей дно, на валики волновой ряби, в углублениях которой собирается терригенный материал. Такие строматолиты в форме рисунка знаков ряби образуют значительные толщи в прибрежной зоне древних морей, образуя при выветривании доскообразные вертикальные стенки строматолитов.
Простую геометрическую классификацию форм протерозойских строматолитов серии Белт без расшифровки ее экологической природы предложили Брайан Логан, Ричард Резак и Гинзбург [36]. Их работа по содержанию аналогична работам И.Н.Крылова, она не несет в себе научного объяснения строматолитовых форм и не представляет большой ценности для экологической морфологии строматолитов. Однако, учитывая сложную природу строматолитовых построек, в соответствии с ботаническим кодексом, названные авторы, совершенно логично, не присваивают выделенным формам, бинарных названий.
Карбонатные наросты цианобактериального происхождения, образуемые бентическими сообществами обрастания на гальках и прибрежных валунах, именуются Nucleella и Nubecularites [14]. Самостоятельные мелкие колонии цианобактерий разных симбиотических образований в прибрежных условиях непрерывного перекатывания волнами образуют онколиты типа Osagia и Ottonosia Twenhofel или Asterospheroides Zhuravleva. Отличие строматолитов и онколитов заключается только в масштабе колоний, как у тубистромий и альтерогофитонов. Как только рост колонии достигает такого размера, что волны уже не в силах перекатывать желвак по дну, последний прирастает и становится новой точкой роста для какой-то формы строматолита. Критерием такого перехода из группы онколитов в строматолиты является величина гидравлического радиуса. В некоторых осадочных толщах, где наблюдаются подобные переходы, критический размер онколитов можно использовать для характеристики бассейна седиментации и определения его гидродинамики и глубины. Скажите мне, где и кто из строматолитчиков это сделал?– никто! — беспросветный эмпиризм, вместо хорошей теории – вот какие бытовали предпочтения в нашей науке. Не зря С.П.Капица в одном из последних номеров переводимого им журнала «Сайентифик Америкэн» за 1983 г. поместил статью о расцвете лженауки в СССР.
- При описании и определении строматолитов В.П.Маслов предлагал четко разделять ископаемую и прижизненную форму, т.к. только прижизненная форма имеет биологический адаптивный смысл и позволяет рационально объяснить возникновение разных форм строматолитов. На материале изучения прижизненных форм можно ожидать создание естественной классификации строматолитов. Ископаемая форма имеет другое содержание и скорее литологический аспект изучения и классификации и, как теперь становится ясным, не может подпадать под линнеевскую системетику и номенклатуру.
Формы строматолитов, вслед за Юлиусом Пиа [37], предлагается рассматривать как формы роста цианобактериальных колоний. А экологически и морфологически сходные формы роста колоний надо рассматривать как жизненные форм (ЖФ) (23,29) и применять к ним эколого-морфологическую классификацию, в общих чертах приведенную здесь. Дело в том, что система жизненных форм адекватно отражает все морфологические особенности строматолитов и является основой для построения их естественной экологической классификации аналогичной линнеевской системе видов организмов [25,40]. Каждая ЖФ как морфологическая единица экологической системы связана с определенными условиями существования или произрастания водорослево-бактериального сообщества, и форма ее отражает весь комплекс действующих на нее факторов морфогенеза. Опираясь на эту теоретическую основу, и учитывая безразличие палеонтологов-строматолитчиков к выделению прижизненой и ископаемой формы у строматолитов, я предлагаю рассматривать «виды и роды» И.Н.Крылова как чисто морфографические искусственные и потому палеонтологически не валидные, как предлагал это сделать А.Г.Вологдин еще много-много лет назад [3]. Уже доказано, что видоспецифических форм строматолитов не существует, почти все формы строматолитов создаются полифитным комплексом цианобактерий и бактерий-симбионтов, сожительствующих с ними в колониях и ценозах.
- Эволюция ЖФ строматолитов шла от простых «аморфных» пленок или матов на дне водоема (экоморфотипы Irregularia, Stratifera, Gongilina и Rifelina) по пути усложнения и усиления роли составляющей биологического морфопоэза, творения собственных адаптивных идиоморфных колониальных форм (экоморфотипы Collenia, Fossella, Conophyton, Osagia и др.). Выявление на материале строматолитовых построек действительного характера подобного биологического морфопроцесса позволяет придать этой классификации эволюционный аспект, аналогичный классификациям растений И.Г.Серебрякова [20], Малькольма Ханзена [35] или Ю.Г.Алеева [1].
Учение о жизненных формах Александра Гумбольдта [6] более широко описывает эволюционный процесс, идущий в живой природе, особенно морфологическое выражение адаптаций к среде, и поэтому способно адекватно отразить и эволюцию строматолитов. Оно без всяких натяжек и попыток подвести их под линнеевскую (видоспецифическую) систематику с ее бинарной номенклатурой способно адекватно описать весь мир строматолитов.
Современные микробиологические исследования цианобактериальных матов в морских и в лагунных мелководьях подтверждают сложную полифитную структуру таких образований и исключают применения искусственной (или видоспецифической) систематики, принятой исследователями строматолитов. В новых работах микробиологи также отдают предпочтение экологической системе жизненных форм [33], пропагандируемую мной с 60-х годов прошлого века [23]. Учитывая всю сумму знаний о строматолитах [16,19,25,29], мы должны ясно сознавать, что морфология построек имеет сугубо экологическую адаптивную природу и вклад генетической конституции самих организмов- строителей в морфогенез и конструкцию постройки минимален и форма их часто вообще не поддается толкованию с позиций наследственной генной морфологии. Формы их могут быть адекватно интерпретированы как формы роста или жизненные формы колониальных цианобактерий или их ценозов — матов [20,25]. Никаких стабильных закрепленных наследственно форм цианобактериальные сообщества не строят, и линнеевская номенклатура, созданная для генетически обусловленных форм, тут явно не уместна [16]. Описываемые на основе внешних различий разновидности строматолитов не могут сравниваться по статусу даже с «распыленными» видами – жорданонами [31], как пытался трактовать их И.Н.Крылов. Строматолитовые формы ближе всего к понятию «жизненная форма» и описываться должны не формально, а в рамках экологическо-морфологического учения о жизненных формах [6,29,40]. Это значит, что выделяемые морфологические единицы должны иметь экологическое обоснование, и генезис их объясняться определенными гидродинамическими, седиментационными и гидрохимическими факторами среды, но сами строматолиты должны рассматриваться как производные органического мира, как вслед за В.П.Масловым, хотя и не совсем удачно, пытался делать И.Н.Крылов. Учение о жизненных формах позволяет адекватно диагностировать и описывать все формы строматолитов и на правильной научной основе завершить начатое им нужное дело морфологической систематизации строматолитов.
- Надо признать, что стратиграфическое значение имеют как сами ЖФ строматолитов (выступающие как маркирующие горизонты в осадочных толщах фанерозоя и рифея), так и составляющие их водорослево-бактериальные комплексы видов, выделяемые с помощью ЭМ в микроструктуре строматолитовых слоев. Эти комплексы видов цианофицей, описываемые независимо от строматолитовых форм [25], анализируются методами обычной биостратиграфии и на их основе формируются списки зональных видов [26,гл.4]. Спектры ЖФ строматолитов характеризуют богатство фациальной картины (экологического ландшафта) стратиграфических подразделений различных рангов и являются хорошими индикаторами фаций осадконакопления. Биогермные ряды, придуманные ad hoc И.Н.Крыловым могут выявить только мелкие экоморфы. Сходство строматолитовых комплексов рифея из различных регионов Земного шара, толкуемое строматолитчиками как эволюционное, скорее объясняется сходством экологии и подобием рифейских фаций, что связано с их гипертрофией с превращением в нимии (термин Наливкина,1952). Гипертрофированные фации, покрывающие громадные пространства однообразных рифейских морей обеспечивают сходство форм строматолитов, но не доказывают одинаковость их стратиграфического положения в разрезе. Без специальных эколого-морфологических исследований и без выявления самих организмов строматолитообразователей на каждом уровне стратиграфического разреза доказать синхронность их невозможно.
Список литературы:
- Алеев Ю.Г.,1986 Экоморфология. Изд. «Наукова Думка».
- Беклемишев В.Н.,1964 Основы сравнительной анатомии беспозвоночных. Том1, Проморфология. Изд. Наука.
- Вологдин А.Г., 1962. Древнейшие водоросли СССР. Изд АН СССР, М.
- Вологдин А.Г., 1963. Строматолиты и фототропизм. ДАН СССР.
- Власов Ф.Я., 1970. Анатомия и морфология строматолитов раннего и среднего протерозоя Южного Урала.// Материалы к палеонт. Урала. Свердловск, УФ АН СССР.
- Гумбольдт Александр фон.,1808. Мысли о физиономике растений. В кн. Картины природы. Изд. Географгиз, М. 1959, с 106-136.
- Гусев А.К., Богатырев В.В., Игонин В.М., Солодухо М.Г. Стратиграфия верхнепалеозойских отложений Актюбинского Приуралья. Изд.КГУ, 1968, 120 с.
- Гурвич А.Г. Теория биологического поля. «Советская наука», 1944, 156 с.
- Давиташвили Л.Ш. В.О.Ковалевский. Изд. АН СССР, 1951, сс.191-212.
- Еленкин А.А.,1907. Орто- и плагиотропный рост с биомеханической точки зрения у лишайников и некоторых других споровых.// Бот.журн. СПб. Общ. Ест. 2,
- Еленкин А.А., Синезеленые водоросли СССР. Т.1-3, 1938- 1946.
- Заварзин Г.А. 1984. Бактерии и состав атмосферы. Изд. Наука, с.192.
- Калиненко В.О. Роль бактерий в формировании железо-марганцевых конкреций.// Микробиология. т..XV, вып.5, 1946.
- Крылов И.Н.,1961 Столбчатые ветвящиеся строматолиты рифейских отложений Южного Урала. Изд. Наука.
- Крылов И.Н., 1975. Строматолиты рифея и фанерозоя СССР. Наука, 243с.
- Маслов В.П. 1960 Строматолиты. Изд. Наука.
- Наумов Д.В., Кин-сун и Хуан Мин-сянь. Главнейшие типы коралловых рифов острова Хайнань. Oceanologia et Limnologia Sinica V.3, №3, 1960.
- Преображенский Б.В. Морфология и палеоэкология табулятоморфных кораллов. Изд. Наука, М.1982.
- Раабен М.Е. Строматолиты.// Бактериальная палеонтология. ПИН РАН, 2002, с. 53.
- Серебряков И.Г.,1962 Экологическая морфология растений. Изд «Высш. Школа» Москва.
- Синнот Э., 1963 Морфогенез растений. Изд. ИЛ,
- Сонин Г.В., 1964 Некоторые типы пермских строматолитов и условия их образования. Итог. науч. аспир.конф. за 1963 г. Изд. КГУ, Казань, с.110-113.
- Сонин Г.В., 1966 Экологическая морфология строматолитов.//Сб. науч. аспир. конф.за 1965 г. Изд Казанск. ун-та,с.28-30.
- Сонин Г.В.,1971 Жизненные формы строматолитов и фации.// Сб. тр.молод. ученых г.Казани. Изд. Обкома ЛКСМ, Казань. с.350-352.
- Сонин Г.В.,1972 Пермские водоросли и строматолиты и их стратиграфическое значение. Диссертация на степень канд. геол.-минер.наук, Казань,172 с., 16 табл. и профиль размещения строматолитов и фаций.
- Сонин Г.В.,1998 Пермские водоросли Актюбинского Приуралья. Изд. Казанск. ун-та, 71с., табл. I-XIV.
- Сонин Г.В.,2007 Фациальные условия распространения и экологический морфогенез пермских строматолитов.//Верхний палеозой России: Стратиграфия и палеогеография. Мат. Всероссийской научн. конф. Изд. Казанск. ун-та. Казань, с.306-309.
- Сонин Г.В., 2011 Диагенетическая микрозональность Перфильева как причина возникновения слоистости в карбонатных строматолитах.//Концептуальные проблемы литологических исследований в России. Мат.6-го Росс. литол. совещ., т.2, с.282-285, Казань.
- Сонин Г.В., 2013, Морфогенез жизненных форм и эколого-морфологическая классификация строматолитов.// Матер. I-й палеоальгол. конф. М..ПИН РАН, с.123-124.
- Сонин Г.В., КалининаТ.А.,Осин Ю.Н., Воробьев В.В., 2015. Стеночные строматолиты соликамского горизонта Пермского Приуралья и их происхождение. // Сб науч. работ «Современные концепции научных исследований», №3(12), ч.8, Москва, с.141-147.
- Тимирязев К.А., 1942, Исторический метод в биологии. Изд.АН СССР, с.81.
- Уоддингтон К.Х. Морфогенез и генетика. М.Мир,1968.
- Штина Э.А., Голлербах М.М. Экология почвенных водорослей.М. Наука,1976, с.143.
- Яковлев Н.Н.,1964 Организм и среда. Наука, с.42-50.
- Hansen M. 1956. Life forms as age indicators. Kopenhagen.
- Logan B.W., Rezak R. and Ginsburg R.N., 1964. Classification and environmental significance of algal stromatolites. -J.Geol.V.72, №1, p.68-83.
- Pia J. in Hirmer. 1927 Handbuch der Palaeobotanik. Bd.1,
- Oliver W.A. Jr. Some aspects of colony development in corals.- J. Paleontol., 1968, v. 42, №5б, p.16-32.
- Shinn E. 1963 Spur and groove formation on the Florida Reef Tract.//J.Sed. Petrol. V.35, №2.p. 291-303.
- Sonin G.V. 1992, The Сyanophyta from Permian red-bed deposites of S.Ural and their significance to solving the morphological and classification problems of stromatolites.//The 29-th Intern.Geol.Cong., Abstracts. Kyoto, Japan.
- Richard Fortey, 2000. Trilobite. Eyewitness to Evolution.
- Fenton C.L.,Fenton M.A. Algae and algal beds in the Belt series of Glacier Nacional Park.- J. Geol.V.39, № 7.
Иллюстрации.
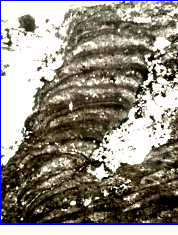
надо рис №3
Рис.1.Связь роста и формы у цианово-бактериальных сообществ. Колонка строматолита растет с наклоном вправо– против течения, тогда как отдельные нити в слоях наклонены влево- по течению. Это совершенно недвусмысленно демонстрирует наличие собственного морфогенетического поля колонии как надорганизменной целостностной системы [25].

Рис.2. Закон геометрического отбора и его применение для объяснения выживания почковидных колоний. Оставить2

надо рис.№5
Рис.5. Параллельные колонки строматолита Gantifera granulose из соликамского горизонта Приуралья. Их образование связано с разделением единой поверхности цианобактериального мата на отдельные бугорки (ценоплазы Твенхофела). Сползающая с бугорков после сезона вегетации отмирающая слизь и химический осадок кристаллов дрюита в многолетнем разрезе формирует отдельные столбики [30], разделяемые инородным веществом био- и хемогенного происхождения. Обычно столбики строматолитов разделяются терригенным или биокластическим веществом. Без этих факторов формирование колонок не наблюдается.

Рис.4. Строматолит Tubistromia, (вверху) и Alterogophyton (внизу) образованные вокруг обломков веток и ствола ископаемого дерева нитчатыми цианофицеями рода Tabantalia из прибрежных фаций тукетской свиты татарского яруса красноцветной перми Актюбинского Приуралья. Рельеф внутреннего канала отражает мелкобугристый рельеф коры дерева и его сучков.
так и остается рис4.
надо рис. 4

Рис.4. Рифельная поверхность пластовых строматолитов из актастинской пачки кунгурского яруса с реки Актасты, Актюбинское Приуралье.
Рис.1. Формы строматолитов актастинской пачки известняков в основании кунгурского яруса Актюбинского Приуралья. Показано как в зависимости от фациальных условий в бассейне формы строматолитов изменяются, хотя все они образованы одним и тем же сообществом цианобактерий [24,25] и трактуются здесь как жизненные формы этого сообщества. Длина профиля с севера на юг около 160 км.

Надо рис № 1[schema type=»book» name=»МОРФОГЕНЕЗ ЖИЗНЕННЫХ ФОРМ И ЭКОЛОГО-МОРФОЛОГИЧЕСКАЯ КЛАССИФИКАЦИЯ СТРОМАТОЛИТОВ» description=»Обсуждается эколого-морфологическая программа исследования форм строматолитов. Приводятся доказательства адекватности применения учения о жизненных формах для описания и систематики строматолитов. Строматолиты рассматриваются как формы роста водорослево-бактериальных симбиотических колоний, как целостных систем надорганизменного уровня организации, обладающих собственным морфогенетическим полем. Приводятся результаты исследования поведения строматолитов в пласте на примере кунгурских строматолитов Южного Урала в рамках предлагаемой программы.» author=»Сонин Геннадий Владимирович» publisher=»БАСАРАНОВИЧ ЕКАТЕРИНА» pubdate=»2017-03-06″ edition=»ЕВРАЗИЙСКИЙ СОЮЗ УЧЕНЫХ_27.06.2015_06(15)» ebook=»yes» ]