
Дальневосточная ветвь бореального биоклиматического экотона Евразии охватывает горно-долинные системы Нижнего Приамурья. Структурное «ядро» этой части бореального экотона представлено подтаежной зоной. Здесь на территории Государственного природного заповедника «Комсомольский» в 2011 и 2012 гг. нами была проведена крупномасштабная ландшафтно-экологическая съемка по методике, изложенной в [4]. На 40 пробных площадях полигона была снята информация по 76 характеристикам состояний лесных экосистем. Основным объектом исследования стали микроландшафты, флористические фратрии и группы типов леса как совокупность древесных пород, представляющая собой, согласно [2], определенный этап лесообразовательного процесса, свойственного данным условиям лесопроизрастания. Флористические фратрии леса [5] были объединены в 6 групп: 1 и 2) маньчжурские мезофильную (Мм) и ксерофильную (Мк); 3) маньчжурско-охотскую (МО); 4) охотскую (О); 5) маньчжурско-ангаридскую (МА); 6 ) ангаридскую (А). Первая, вторая, четвертая и шестая фратрии являются базовыми и они широко известны [3]. Третья и пятая фратрии – буферные (смашанные), они выделены нами. Рассматриваются также 6 групп типов леса: 1) широколиственные леса (Шл); 2) кедрово-широколиственные (КШ); 3) елово-широколиственные (ЕШ); 4) пихтово-еловые (ПЕ); 5) лиственничные леса (Лист); 6) лесо-болотные комплексы, лиственничные мари (ЛБ).
Микроландшафты, по [1], выделены по сочетанию двух геоморфологических признаков групп лесных биогеоценозов – типов их мезомасштабных местоположений и солярной экспозиции склонов (для транзитных типов МП). Эти объединения характеризуют сложную горно-долинную систему мезомасштабных ландшафтных сопряжений (мезокатен). Выделены семь микроландшафтов: 1) горные элювиально-трансэлювиальные (ТЭГ) сублитоморфные; 2) горные транзитные теневых склонов (ТГТ) лито-гидроморфные; 3) горные транзитные солнцепечных и нейтральных склонов (ТГС) литоморфные; 4) предгорные трансаккумулятивные (ТА) сублитоморфные; 5) долинные элювиальные (ЭД) сублитоморфные; 6) долинные аккумулятивные (АД) субгидроморфные; 7) долинные супекракавальные (Saq) гидроморфные. Были выделены следующие лесные группы: 1) широколиственная (Шл); 2) кедрово-широколиственная (КШ; 3) елово-широколиственная (ЕШ); 4) пихтово-еловая (ПЕ); 5) лиственничная (Лист); 6) лесо-болотная (ЛБ), представленная лиственничными марями. Первая и вторая группы отнесены, согласно [7], к маньчжурскими флористическим фратриям (мезофильной и ксерофильной), а четвертая и пятая – соответственно к охотской и ангаридской фратриям. Буферную маньчжурско-охотскую фратрию представляет группа елово-широколиственных лесов.
Полученный материал был проанализирован с помощью методов теории информации [6]. В данной статье представлены результаты бинарной ординации с использованием частного коэффициента связи Сij отдельных градаций (состояний) явления (ai) и фактора (bj). С помощью бинарной ординации устанавливали систему экологических ниш каждого значения (градации) явления А в пространстве значений фактора В. По системе экологических ниш вычислялась таксономическая норма того или иного функционального признака для каждой группы типов леса. Значения параметра Сij вводились в расчеты в качестве «весовых» коэффициентов. По матрице значимых величин Сij > 1 строились соответствующие графики, причем градации явления располагались в порядке роста, либо снижения их экологическим доминантов по градациям данного фактора. Через доминанты затем проводилась огибающая кривая, дающая достаточно четкое представление о наиболее существенной тенденции изучаемой связи. Многие характеристики лесных экосистем имеют не только основную экологическую нишу по тому или иному фактору (с соответствующим экологическим доминантом и с «размытой» частью ниши), но и своего рода анклав, отделенный на графике частных коэффициентов связи от самой ниши более чем на одну градацию фактора.
Модельная территория Комсомольского заповедника характеризует только низкогорно-равнинную часть Приамурского бореального экотона (с абс. высотами ниже 1000 м). Как известно, в низкогорьях и предгорьях уже проявляются признаки высотной биоклиматической поясности, однако эта картина существенно нарушается другими морфо-климатическими факторами субрегионального и локального порядка – прежде всего, циркуляционной и солярной экспозицией склонов, а также барьерными эффектами и климатической инверсией в условиях горно-долинного рельефа. Фито-экологическое и ландшафтообразующее влияние этих факторов вполне отчетливо проявляется на территории заповедника, что отражено в общей тесноте информационных связей лесного покрова с различными геоморфологическими характеристиками. Если мера сопряженности К(А;В) флористических фратрий и типов леса с абсолютной высотой равна соответственно 0,126 и 0,102, то по солярной экспозиции склонов она оказывается еще выше – 0,174 и 0,157. Существенную роли играют также крутизна склона (К(А;В) = 0,116–0,118) и литологический состав коренных пород (К(А;В) равен соответственно 0,117 и 0,270).
В горных странах зональность проявляется гораздо более разнообразно, чем на равнинах, что отразилось в дальнейшем развитии докучаевской теории географической зональности. Показано, что суперпозиция двух начал региональной дифференциации горной территории: морфотектонического и макроклиматического, – предопределяет целый спектр зональных явлений: широтного, высотного, продольно- и экспозиционно-циркуляционного, барьерного, солярно-экспозиционного. Сочетаясь, они дают сложные формы так называемой дислокационной зональности. В этой концепции зональность тесно связывается с понятием вектора (градиента) и определяется как закономерная, упорядоченная в пространстве направленность изменений свойств природных комплексов в целом и их отдельных компонентов.
Проследим, насколько проявляется дислокационная зональность в низкогорьях Приамурского бореального экотона. В качестве локальных фиоценологических индикаторов этой зональности будут выступать флористические фратрии и типы леса, а комплексным субрегиональным индикатором станут микроландшафты Комсомольского заповедника. С этой целью представим ороклиматическую систему данной территории как суперпозицию двух ведущих в условиях гор абиотических факторов – абсолютной высоты (А) и экспозиции склонов (В) (Табл.).
Таблица. Ординация точек по степеням абсолютной высоты (А) и солярно-циркуляционной экспозиции (В).
|
Абс. высота, м. (тип мезоместоположения) |
Экспозиция |
|
| В1 – СЗ-С-СВ-В | В2 – ЮВ-Ю-ЮЗ-З | |
| А1 – 20–110 (Saq, Адол, Эдол) | Градация 1 | Градация 3 |
| А2 – 110–310 (Т гсолн, ТА предгорн.) | Градация 4 | Градация 5 |
| А3 – 310–680 (Т гтен, Э-ТЭ г |
Градация 6 |
Градация 7 |
| А4 – нижний ярус | В0 Градация 2 | |
Бинарная ординация указанных фитоценологических и ландшафтных индикаторов по фактору дислокационной зональности (рис. 1), с достаточно высокими коэффициентами сопряженности, убедительно свидетельствует о том, что в условиях муссонно-континентального климата высотная биоклиматическая поясность низкогорья проявляется весьма избирательно и в целом существенно нарушается экспозиционными контрастами. Преломление высотной поясности комплексным фактором циркуляционно-солярной экспозиции обусловлено, как можно полагать, двойственностью самой региональной биоклиматической системы Юга Дальнего Востока. Эта двойственность состоит в соразмерном выражении широтной зональности субокеанического типа (широколиственные леса – смешанные леса – темнохвойные леса) и долготной секторности восточного окраинно-материкового типа. Роль секторности проявляется прежде всего во внедрение по долинам рек и С3–С–СВ–В горным склонам средней полосы соответственно равнинных и горных лиственничных лесов – представителей мерзлотно-таежной восточносибирской флоры. В этой же полосе и на склонах этих же экспозиций в результате смешения последней с ксерофильной маньчжурской флорой возникли сложные лиственничники, с участием широколиственных пород, относящиеся уже к буферной маньчжурско-ангаридской фратрии.
Отсутствует также однозначная высотно-поясная приуроченность самих маньчжурских флористических фратрий как основных фитоценологических представителей Приморско-Приамурской субпацифики. Широколиственные и кедрово-широколиственные леса встречаются почти по всему высотному профилю низкогорья и к тому же имеют два экологических доминанта: 1) ксерофитный вариант на равнинах и в предгорьях и 2) вариант мезофитный – на южных и юго-западных склонах в верхней полосе хребтов. В этой же полосе, но на склонах северо-восточной экспозиции появляется доминирующая область пихто-ельников – следующего по высоте типа леса, представленного охотской флорой. Как видим, высотная поясность здесь хотя и начинает проявляться, но весьма избирательно – по склонам наиболее благоприятной для этого экспозиции, в соответствии с известным «правилом ландшафтного предварения» Вальтера-Алехина.
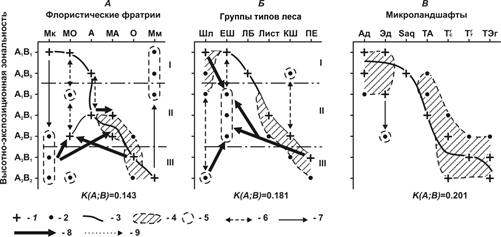
Рис. 1. Распределение лесных биогеоценозов в пространстве высотно-экспозиционной зональности.
Трансгрессия по солнцепечным склонам с верхних склонов гор в среднюю полосу Мк и ОК флор и их смешение привело к формированию елово-широколиственных лесов. Однако леса этой же буферной фратрии столь же широко распространены в предгорьях, с анклавом на южных склонах нижнего пояса гор.
Рассмотрим более подробно высотно-зональное распределение лесного покрова на территории Комсомольского заповедника (рис. 2). Большинство флористических фратрий имеют как основную, так и второстепенную экологические ниши, либо определенные анклавы по фактору абсолютной высоты. Хотя экологические доминанты выражены достаточно четко (с вероятностью до 40–45%), сами флористические ниши весьма «размыты», что указывает на существенное «растекание» базовых и буферных лесных флор от своих очагов под влиянием локальных геоморфологических факторов, которые способны обеспечить им повышенную устойчивость к фоновым климатическим сигналам. Вместе с тем, типы леса имеют весьма суженные экологические ниши, часто вообще без «размытой» части (см. рис. 2 Б). Это свидетельствует об уже устоявшейся высотной дифференциации типов леса, но одновременно – о повышенной их чувствительности к климатическим колебаниям.
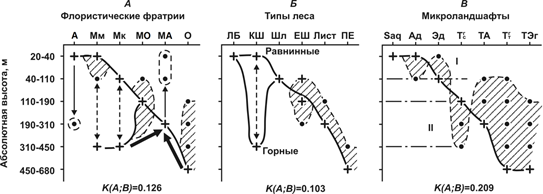
Рис. 2. Распределение лесных биогеоценозов в зависимости от абсолютной высот местности.
Мезофильные маньчжурские леса имеют две достаточно оторванных друг от друга группы ареалов: 1) на равнинах и в предгорьях и 2) в средней полосе низкогорных хребтов на отметках 300-450 м. В первом случае это почти на 80% чисто широколиственные леса, которые приурочены к хорошо и избыточно увлажненным супераквальным локальным субгоризонтальным местоположениям на надпойменных террасах Амура и днищах малых речных долин, с уклонами не более 5о. Вторую группу Мм ареалов образуют кедрово-широколиственные леса, занимающие главным образом элювиальные и трансэлювиальные местоположения на гребнях хребтов и привершинных солнцепечных и нейтральных склонах наиболее высокой крутизны (до 20о и более). Данный факт подчеркивает известное положение о том [2], что кедровники юга Дальнего Востока – это, прежде всего, горные леса.
На предгорных равнинах и в речных долинах широко распространены также леса ангаридской флоры, резко контрастирующей по своим эколого-физиологическим характеристикам с флорой маньчжурской. Как видим, арена локальных контактов основных фитоценологических представителей субпацифики и палеопацифики приурочена на Приамурском бореальном экотоне к низменным равнинам и предгорьям, причем оба они встречаются преимущественно на субгоризонтальных поверхностях. На горных склонах их экологические доминанты разнесены по разным солярным экспозициям: солнцепечной и нейтральной для широколиственных лесов маньчжурской мезофитной флористической фратрии и теневой – для лиственничников фратрии ангаридской.
Маньчжурские ксерофитные леса имеют так же две четко выраженные высотные доминирующие области. Первая приходится на нижнюю полосу низкогорий и приурочена исключительно к нейтральным склонам. Вторая область занимает солнцепечные и наиболее крутые (15-20о) горные склоны на отметках высот 300–450 м. Пихтово-еловые и елово-широколиственные леса охотской флоры занимают верхнюю полосу (450-700 м.) – гребни хребтов и привершинные крутые склоны, как правило, теневые и средней крутизны (не более 10-15о). Резко выраженный экологический доминант основной ниши этих лесов и отсутствие ее «размытой» части указывают на то, что «очаг» формирования аянских хвойных лесов в Нижнем Приамурье достаточно сильно связан с биоклиматическими условиями теневых склонов верхней полосы низкогорий, причем их пространственная ниша оказывается очень узкой, приуроченной главным образом к ТА и А локальным местоположениям. Наконец, в средней наиболее широкой полосе, господствуют буферные флоры, производные от маньчжурской – МО и МА.
Флористические фратрии средней и верхней полосы (О, МО и МА) имеют анклавы, расположенные ниже по склонам от их основных высотных ниш и указывающие таким образом на нисходящую локализованную трансгрессию двух базовых флор – маньчжурской мезофильной и охотской психрофильной.
Доминанты двух буферных флористических фратрий лесов: маньчжурско-охотской и маньчжурско-ангаридской, — сформировались в территориально близких друг к другу высотных полосах. Смешение охотской флоры с маньчжурской шло от верхней полосы низкогорий, где находится «очаг»-доминант аянских пихто-ельников, вниз по склонам – навстречу доминанту экологической ниши горных мезофильных кедрово-широколиственных лесов. Формирование елово-широколиственных лесов, т.е. внедрение аянской ели в ясенево-дубово-липовые сообщества происходило главным образом на нейтральных и реже теневых трансэлювиальных склонах, куда и сместился экологический доминант этой буферной МО фратрии. Кроме того, ее доминант оказался на 100-200 м. ниже по отношению к доминанту горной Мм фратрии. Возможно, это является свидетельством сверхвекового снижения летних температур и роста годовой увлажненности климата. Если такое предположение верно, то и общую картину трансгрессии «размытого» поля экологических ниш большинства флористических фратрий, с образование анклавов, можно объяснить именно этим сверхвековым климатическим трендом.
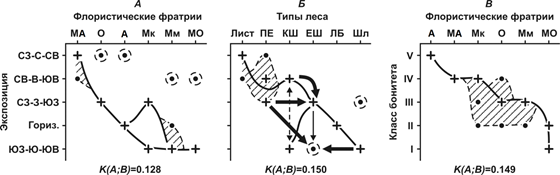
Рис. 3. Распределение лесных биогеоценозов в зависимости от экспозиции склона.
По солярно-циркуляционной экспозиции (рис. 3) хорошо дифференцируются ЕШ и ПЕ леса, с одной стороны, и КШ – с другой. Аянские пихто-ельники широко распространены на всех нейтральных склонах и охватывают также склоны северной ориентации. Однако экологический доминант лесов О флоры приходится на склоны западных румбов, обращенные к влагонесущим воздушным массам. В то же время область доминирования широколиственных лесов базовой Мк флоры приходится на склоны южных румбов и восточные. Трансгрессия горных широколиственных лесов с восточных склонов в пихто-ельники западных склонов привела к формированию на этих склонах буферной МО формации – елово-широколиственных лесов. Со своей стороны, распространение пихто-ельников вниз по долинам рек и внедрение их в Мм широколиственные леса привели к формированию долинных анклавов этой буферной фратрии.
Итак, два противоположных «полюса» флор: Мм и А фратрии имеют свои области доминирования на почти одинаковых абс. высотах, в условиях близкой солярной экспозиции и при одинаковом равнинном мезо- и микрорельефе. Буферная между ним флора (МА) распространена в основном на Э и ТЭ типах локальных МП, как на долинных плакорах, так на и гребнях и вершинах хребтов. Здесь в обоих случаях идет смешение маньчжурской мезофильной и
ангаридской континентальной флор, с внедрением лиственницы в кедрово-широколиственные леса. Однако здесь же распространены ишироколиственные леса с дубом (Мк флора) – следствие одновременной аридизации мезофильных маньчжурских лесов. Как видим, реально существующая мозаика рассматриваемых флор в Нижнеамурской низкогорной субпацифике не привязана жестко к геоморфологическому каркасу и сформирована собственной биоклиматической системой – вековыми и сверхвековыми климатическими колебаниями, а также эктогенетическими сукцессиями самой лесной растительности.
Таким образом, в результате различного сочетания геолого-геоморфологических факторов и в связи с геоэкотонным положением заповедника здесь наряду с «коренными» образовались смешанные лесные сообщества. На локальном уровне это выражается в разнообразной «мозаике» лесных биогеоценозов, представленных различными типами леса и микроландшафтами.
СПИСОК ЛИТЕРАТУРЫ
- Глазовская М. А. Геохимические основы типологии и методики исследований природных ландшафтов. М.: Изд-во МГУ, 1964. 230 с.
- Колесников Б. П. Кедровые леса Дальнего Востока. Труды ДВ филиала АН СССР, 2 (4). М.-Л., Изд-во АН СССР, 1956. Т. 2 (4), сер. ботан. 262 с.
- Колесников Б. П. Высокогорная растительность Среднего Сихотэ-Алиня. Владивосток: Дальневост. Кн. Изд-во, 1969. 106 с.
- Коломыц Э. Г. Локальные механизмы глобальных изменений природных экосистем. М.: Наука, 2008. 427 с.
- Куренцова Г. Э. Естественные и антропогенные смены растительности Приморья и Южного Приамурья. Новосибирск: Наука, 1973. 231 с.
- Пузаченко Ю. Г, Скулкин В. С. Структура растительности лесной зоны СССР. Системный анализ. М.: Наука, 1981. 275 с.
- Сочава В. Б. Геотопология как раздел учения о геосистемах // Топологические аспекты учения о геосистемах. Новосибирск: Наука, 1974. С. 3–86.[schema type=»book» name=»ЭМПИРИКО-СТАТИСТИЧЕСКИЕ МОДЕЛИ МОНОСИСТЕМНОЙ ОРГАНИЗАЦИИ ЛЕСНЫХ СООБЩЕСТВ НА БОРЕАЛЬНОМ ЭКОТОНЕ НИЖНЕГО ПРИАМУРЬЯ» description=»На основе проведенного в 2011-2012 гг. на территории Комсомольского заповедника ландшафтно-экологического исследования выполнен информационный анализ межкомпонентных ландшафтных связей в лесных природных комплексах Нижнего Приамурья. Построена генеральная информационно-статистическая модель и парциальные модели ландшафтно-экологических связей, отражающие основные закономерности распределения лесных сообществ на исследуемой территории. » author=»Петренко Полина Сергеевна, Коломыц Эрланд Георгиевич» publisher=»БАСАРАНОВИЧ ЕКАТЕРИНА» pubdate=»2017-02-02″ edition=»ЕВРАЗИЙСКИЙ СОЮЗ УЧЕНЫХ_26.09.15_10(18)» ebook=»yes» ]