
Флуктуации химических реакций, белков и функций живого организма подробно описаны в книге «Molecular science of fluctuations toward biological functions» (2016) [14] /с акцентом на термодинамические флуктуации/, в работах д.б.н. С.Э. Шноля и сотр. [5, 6] /с акцентом на макрофизическую причину флуктуаций/, Ю.Ф. Крупянского и сотр. [3] /с акцентом на кинетическую теорию/.
В биомолекулярных системах со многими степенями свободы, включающих белки и нуклеиновые кислоты, существует астрономическое число состояний (структурная динамика). [11] Конформация биомолекул меняется при сдвигах рН (разворот гемагглютинина вируса гриппа при адгезии к клеткам-мишеням …), ионной силы, при фосфорилировании, ацетилировании (ацетилирование гистонов /Рис. 1/), взаимодействии рецептора с лигандом, антитела с антигеном, при работе лейциновой застежки-молнии, цинковых пальцев, кластеризации рецепторов и пр. Данные изменения влияют и на флуктуационные характеристики.
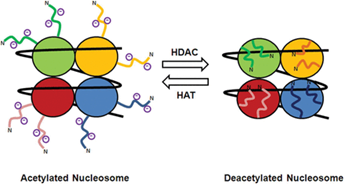
Рис. 1. Изменение заряда (и конформации ) гистоновых хвостов при
ацетилировании [по10]. Характер перестроек определяет
гистоновый «код».
Согласно результатам исследований [14], флуктуируют (значимо для следствий функций протеина) белки, выполняющие регуляторную и эффекторную роль; акцентируется внимание на внутреннеприсущую неупорядоченность белков; обсуждаются возможные конформационные состояния и подсостояния, биомолекулярные системы с быстрым конформационным обменом, изменение характера флуктуаций при взаимодействии белковой молекулы с лигандом. Показано усиление флуктуаций белков во время реакций. [13] Выдвигается теория молекулярного распознавания в свете (в аспекте) структурных флуктуаций биомолекул. [14]
Согласно Л.Б. Меклеру [4], многие антигены (эпитопы) имеют форму шпильки (петля и «шейка»); за счет постоянных перемещений (перескоков) диаметр петли меняется. Сдвиги в иммунодоминантной петле белка gp120 вируса иммунодефицита человека (ВИЧ-1),возможно, препятствуют взаимодействию с антителами. [1] По аналогии можно предположить подобную динамику (Рис. 2) стеблей и петель шпилек РНК (полианиона), что дает математическую квантованность состояний (и, возможно, частот изменения заряда петли) – обработку информации /?/. Динамика молекулы РНК описана в работе [9] и др.

Рис. 2. Возможная динамика (скачки) по тандемным палиндромным
повторам в шпильках молекул белков и РНК.
Современное моделирование молекулярной механики (ММ) проводится с привлечением методов квантовой механики (КМ; QM) – методы QM/MM. [12] Последние используются в биологической химии, в частности, при изучении функционалов типа кластеров переходных металлов в металлоферментах для описания поведения в сложных средах. [8]
Экспериментальная часть работы. Цель — исследование флуктуаций времени свертывания (рекальцификации) плазмы крови. Кровь забирали из вены в пробирки с цитратом натрия от лиц 18-35 лет. Получали центрифугированием обогащенную и обедненную тромбоцитами плазму. К 0,1 мл плазмы добавляли 0,2 мл хлористого кальция и измеряли время появления сгустка («время рекальцификации») 20-100 раз подряд (в каждом образце плазмы). Результат. Исследование флуктуаций времени рекальцификации показало, что разброс может быть большим или меньшим у каждого исследуемого лица и даже воспроизводимым по форме гистограммы распределения полученных инвариант (Рис. 3).
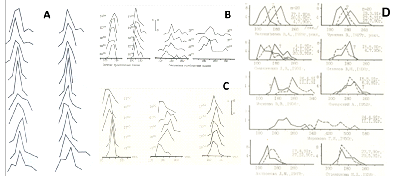
Рис. 3. Некоторая повторяемость формы гистограмм распределений
времени свертывания крови (времени рекальцификации плазмы)
разных доноров (по 20 замеров для каждой гистограммы).
A – десять доноров (попарно слева и справа). В и С – четыре (В)
и три (С) донора (замеры на протяжении нескольких часов (снизу
вверх). D — девять больных шизофренией (замеры дважды с
интервалом несколько месяцев).
Коэффициент парной корреляции различных комбинаций среднеарифметических величин (n=20) времени свертывания составил 0,92, коэффициента вариации — 0,65.
В следующих экспериментах оценивали влияние in vivo адреналина и мединала на флуктуационный разброс времени свертывания крыс. Кровь (примерно 5 мл) забирали у беспородных белых крыс из брюшной аорты в шприц с цитратом натрия, центрифугировали, получали обедненную тромбоцитами плазму. Данного объема плазмы хватало на 18-30 замеров. Опытной серии крыс предварительно в яремную вену вводили раствор адреналина (5 и 0,5 мкг/кг веса тела животного) или мединала (10 мг/кг). Адреналин был выбран в качестве агента, повышающего психическую напряженность, мединал — как обратно действующий успокаивающий препарат. Кровь забирали через 3-5 минут и через час после введения.
Адреналин в дозе 5 мкг/кг резко сужает амплитуду флуктуаций времени рекальцификации (с 13±1% в контроле до 8±1% в опыте); в концентрации 0,5 мкг/кг не оказывал подобного действия. (Рис.4) Мединал, напротив, через 3 минуты после введения расширял (до 18±1,4%) спектр разброса получаемых инвариант; через час данного явления не наблюдалось. (Рис. 4)
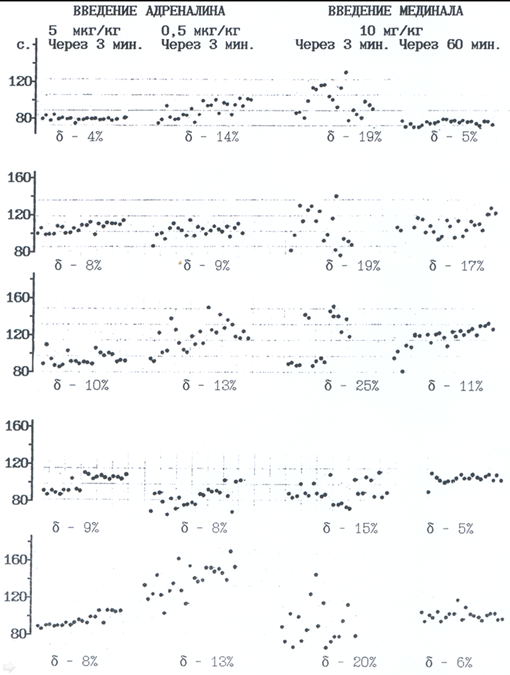
Рис. 4. Влияние адреналина и мединала (внутривенное введение) на флуктуации
времени свертывания плазмы крови крыс. Адреналин (гормон стресса)
снижает показатель вариативности времени свертывания крови
(мобилизует систему); мединал («снотворное») действует обратным
образом.
Исследования «снизу-вверх» (начиная от чистых систем – один объект, один изучаемый фактор) и «сверху-вниз» (in vivo; с множеством возможных эффекторов) относятся к разным сферам науки и данные параллели пока не стыкуются. Тем не менее возможны обобщения (синергетические, философии биологии). Пользуясь тетрадным методологическим подходом, выделим в итоге разные варианты динамики белковых молекул. (Таблица 1)
Таблица 1.
Варианты поведения белковых молекул (тетрадный подход). Наброски.
| Limà0 | Флуктуации | Функция | Структура |
|
Молекулярный уровень [структурный подход] |
|||
| Статика
(белковые полимеры, субмолеку-лярная класте-ризация (амилоиды, капсиды вирусов …)
|
Флуктуационное
поведение [внешнее – внутреннее] — термодинамичес- кая — космофизическая — флуктуации «информационные» (маркерные; возможно, с характером «Я» — эпигенетические, кванты в петлях …) |
Функциональ-ное поведение
(в активном центре эффекторной молекулы) |
Структурное поведение
— надмолекулярная (при взаимодействии с иными лигандами, самосборка при полимеризации …) [внешнее] — молекулярная (меха- нические перемеще- ния /механика/) — субмолекулярная (переходы «альфа- спираль»-шпилька …) [внутреннее] |
|
Каскадные реакции (AàBàCà…) [функциональный подход] — внутриклеточные сигнальные пути — ферментные системы крови (свертывающая, комплемента и др.) |
|||
| Резонанс | Возможно явление
quorumsensing («чувство локтя»; единое коммуника- тивное поведение) — Через сигнальные молекулы в био- пленках микробов |
? «Удвоение» в
динамике (nà 2nà 4nà …) [Количество] Масштабиро-вание в «частотах» ? |
? Кооперация
(агрегация, комплексообразование …) |
|
? Карты торсионной плотности (энергия-ландшафт)* [флуктуационный подход] |
|||
| Стохасти-
ческая синхрони- зация ? |
Стиль
(индивидуальный …) — Модели поведения ферментов — Аттракторы … Клетка [и организм] модулирует энергетические ландшафты [14] |
?
Потоки энергии, кручения энергии …
|
?
Ландшафт динамики белков (молекул) организма, географи-ческого [2] региона – флуктуации белков растений, растущих над месторождением полезных ископаемых … |
| Фликкер-шум (в неравновес-
ной термоди- намике, био- логии …) – фрактальный |
Цветные (и
индивидуальные) шумы |
Коричневый шум
(сдвиговый /«походкой пьяного»/) |
Белый шум-? |
Примечание. * Термины заимствованы из источника [7].
Вывод 1 (по таблице). Флуктуационное поведение молекул
Пронизывает разные уровни организации материи, структурная
динамика – лишь объекты, объединенные данной структурой.
Вывод 2. Между внешним и внутренним – механика (итоговое
передвижение /решение/ по эпигенетическому ландшафту)
[триада – внутреннее, внешнее, итоговое поведение].
Список литературы:
- Андрианов А. Конформационный анализ белков: теория и приложения. – Litres, 2014. – 518 с.
- Баласанян С.Ю. Динамическая геоэлектрика. — Новосибирск: Наука, СО, 1990. — 232 с.
- Крупянский Ю.Ф., Гольданский В.И. Динамические свойства и энергетический ландшафт простых глобулярных белков // Успехи физических наук. – 2002. – Т. 172,N – С. 1247-1269.
- Меклер Л.Б. О специфическом избирательном взаимодействии между аминокислотными остатками пептидных цепей // Биофизика. — 1969. — Т. 14, N — С. 581-584.
- Шноль С.Э. Космофизические факторы в случайных процессах // Stokholm: Svenskafysikarkivat. – 2009. – 388 с. https://digitalphysics.ru/pdf/Kaminskii_A_V/shnoll2009ru.pdf
- Шноль С.Э., Коломбет В. А., Пожарский Э. В. и др. О реализации дискретных состояний в ходе флуктуаций в макроскопических процессах // Успехи физических наук. – 1998. – Т. 168, N – С. 1129-1140.
- Chakrabarti A., Surolia A. (ed.). Biochemical Roles of Eukaryotic Cell Surface Macromolecules. – Springer, 2015. – 411 p.
- Cui Q. Perspective: Quantum mechanical methods in biochemistry and biophysics //The Journal of Chemical Physics. – 2016. – Vol. 145, N 14. – P. 140901.
- Dethoff E.A., Chugh J., Mustoe A.M., Al-Hashimi H. Functional complexity and regulation through RNA dynamics // Nature. – 2012. – Vol. 482, N 7385. — P.322-330.
- Hawtree S., Muthana M., Wilson A.G. The role of histone deacetylases in rheumatoid arthritis fibroblast-like synoviocytes // Biochemical Society transactions. – 2013. – Vol. 41, N 3. – P. 783-788.
- Okamoto Y. Structural Fluctuations of Proteins in Folding and Ligand Docking Studied by Replica-Exchange Simulations // Molecular Science of Fluctuations Toward Biological Functions. – Springer Japan, 2016. – P. 183 — 204. https://scholar.google.ru/citations?view_op=view_citation&hl=ru&user=4XMv5IoAAAAJ&cstart=200&pagesize=100&citation_for_view=4XMv5IoAAAAJ:WwIwg2wKZ0QC
- Shen L., Wu J., Yang W. Multiscale Quantum Mechanics/Molecular Mechanics Simulations with Neural Networks // Journal of Chemical Theory and Computation. – 2016. – Vol. 12 (10). – P. 4934-4946.
- Terazima M. Time-resolved detection of protein fluctuations during reactions // Molecular science of fluctuations toward biological functions. – Springer Japan, 2016. – P. 1-27.
- Terazima M., Kataoka M., Ueoka R., Okamoto, Y. (Eds.). Molecular science of fluctuations toward biological functions. – Springer, 2016. – 269 p.[schema type=»book» name=»ФЛУКТУАЦИИ БЕЛКОВ, В ЧАСТНОСТИ, СВЕРТЫВАЮЩЕЙ СИСТЕМЫ КРОВИ» description=»Структурная динамика многих биомолекул зависит от различных факторов среды, внутренне присущих перестроек (за счет липкости, комплементарности пар нуклеотидов /комбинаторика/ …), кооперации с иными молекулами (часто специфическими) и др.В работе показана вероятность влияния системной характеристики (в какой-то степени индивидуализации /разные люди/) на вариабельность активности белков коагуляционного каскада.» author=» Ложкина Анна Николаевна» publisher=»БАСАРАНОВИЧ ЕКАТЕРИНА» pubdate=»2017-01-17″ edition=»ЕВРАЗИЙСКИЙ СОЮЗ УЧЕНЫХ_30.10.16_31(2)» ebook=»yes» ]