
Диплоидный набор хромосом (2n) является важнейшим критерием при описании вида. У личинок комаров-звонцов (Chironomidae), чьи политенные хромосомы из слюнных желез являются удобным объектом для изучения популяционных явлений и реакции на неблагоприятные условия среды, анализ диплоидного числа, как правило, дает однозначные результаты. Однако, имеется ряд исключений, связанный с особенностями репликации ДНК в политенных хромосомах по сравнению с митотическими. По мере возникновения новых методов исследования генома и публикации новых данных о поведении наследственного материала в ходе онтогенеза, появляется необходимость ревизии 2n у отдельных видов.
Объектом наших исследований послужили три вида из рода Pseudodiamesa Goetgh. – Рseudodiamesa stackelbergi Goetgh. (= Sindiamesa nivosa) (2n=8) [6, 7, 8], Pseudodiamesa ex gr.branickii Nowicki (2n=8) [2, 5], Pseudodiamesa sp. (2n=8) [7, 8] и вида из рода Anatopinia – A. plumipes Fries (2n=6) [9, 8].
Перечисленные виды были изучены кариологически, путем приготовления давленых препаратов политенных хромосом, окрашенных по рутинной ацетоорсеиновой методике. У всех 4-х видов хромосомы объединялись в псевдохромоцентр, который при приготовлении временных давленых препаратов обычно распадался на отдельные плечи, вследствие недорепликации прицентромерного гетерохроматина [1, 3]. При этом хромосомы сохраняли разные эктопические контакты друг с другом, что затрудняет определение принадлежности плеча к той или другой хромосоме. В таких случаях элементы диплоидного набора уточняли по метафазным и анафазным пластинкам из гониальных или ганглиальных клеток.
Изначально мы принимали каждое хромосомное плечо за самостоятельную хромосому, то есть кариотип P. stackeldergi характеризовался 2n=14, Pseudodiamesa sp. 2n=12, A. plumipes — 2n=10. Как показали дальнейшие исследования, такой подход был неверен.
Политенные хромосомы образуются в ходе множественных актов репликации ДНК, однако далеко не весь ядерный материал участвует в этом процессе. Так, у Р. stackelbergi была обнаружена хромосома, представляющая собой бесструктурную гетерохроматиновую массу [6], которая, предположительно, представляла собой недореплицированное плечо малой ядрышкообразующей хромосомы [4]. Таким образом, несоответствие в данных о митотических и мейотических хромосомах, с одной стороны, и политенных — с другой потребовало пересмотра интерпретации ранее полученных данных.
Статистическими методами были проанализированы случаи остаточной конъюгации плеч хромосом в кариотипах вышеуказанных видов после разрушения хромоцентра. Было установлено, что распределение сочетаний неслучайно на 5% уровне значимости. Некоторые сочетания встречались достоверно чаще остальных, с частотами в интервале 12,3 – 33,5%. Для P. stackelbergi таких сочетаний было три, когда объединялись плечи I+IV, II+V и III+VI. Плечо VII оставалось свободнолежащим и преимущественно конъюгировало только с бесструктурной гетерохроматиновой массой, а с остальными плечами – крайне редко и нерегулярно. Таким образом, можно считать, что диплоидный набор у данного вида 2n=8. Хромосомы, которые мы считали ранее
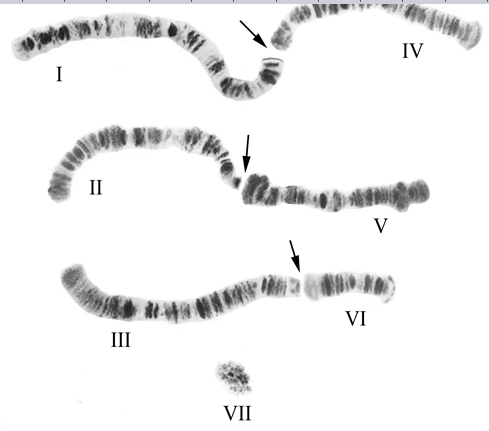
Рис. Кариотип Рseudodiamesa stackelbergi Goetgh. Типичный случай диссоциации хромоцентра на 7 отдельных элементов. Римскими цифрами с I по VII указаны порядковые номера хромосомных плеч, ранее считавшиеся отдельными хромосомами. Стрелками указаны точки разрыва хромосом в пределах хромоцентра и предполагаемое местоположение центромер. Фактически пара I+IV составляет хромосому I, пара II+V – хромосому II, пара III+VI – хромосому III, у свободнолежащей акроцентрической хромосомы VII статус не меняется.
телоцентрическими, в действительности являются двуплечими. Все три пары больших гомологов субметацентрические. Четвертая пара гомологов выглядит как сгусток аморфного гетерохроматина с более темным участком прицентромерного гетерохроматина в центре (Рис) [6].
У Pseudodiamesa ex gr.branickii по разным данным диплоидный набор оценивался как 2n=6 [7, 8]. У этого вида в фолликулах была обнаружена одна малая хромосома, которая не проявляется в клетках слюнных желез, то есть не подвергается политении [2]. Ревизия сочетаний хромосомных плеч показала, что наиболее часто встречаются три устойчивых сочетания с достоверностью 5%.
Однако, принимая во внимание данные Захариаса [2], диплоидное число этого вида равно 8, так как одна пара гомологов недореплицируется в ходе политенизации.
В кариотипе вида Pseudodiamesa sp. 12 хромосомных плеч достоверно образовывали сочетания пар плеч I+IV и II+III, (то есть две субметацентрические пары гомологов), тогда как V и VI пары оставались свободнолежащими и, соответственно, являлись самостоятельными акроцентрическими хромосомами. Таким образом, было установлено, что диплоидный набор вида – 2n=8.
В кариотипе A. plumipes из подсемейства Tanypodinae было обнаружено 10 хромосомных плеч, достоверно образующие два типа устойчивых сочетаний, фактически представляющих собой две пары субметацентрических гомологов (хромосомы I и II). Эти две хромосомы имеют небольшие расширенные зона, а кроме того хромосома I – ядрышковая. Ядрышко расположено рядом с центромерой. Пятая пара хромосом остается свободной, это хромосома III. Однако для акроцентрика эта хромосома относительно более длинная, чем можно наблюдать в более прогрессивном подсемействе Chironominae, и в кариотипах подсемейства Diamesinae. Возможно, этот признак является особенностью кариотипов Tanypodinae. Однако данный тезис еще ждет своего подтверждения, так как политенные хромосомы таниподин являются сложным объектом из-за низкой степени политении и многочисленных межхромосомных контактов в объеме клеточного ядра. Итак, вид A. plumipes характеризуется 2n=6.
Подводя итог, следует отметить необходимость подобных работ для уточнения научных фактов по мере поступления новых данных. В тех случаях, когда виды имеют длительный репродуктивный цикл, малоплодовиты или фенологически однообразны, классическая генетика бессильна. А методы молекулярной генетики не скоро станут достаточно дешевы и просты для решения таких задач.
Работа выполнена при финансовой поддержке Программы фундаментальных исследований Президиума РАН «Биоразнообразие природных систем», подпрограммы «Генофонды живой природы и их сохранение».
Список литературы
- Bauer H. Beitrage zur vergleichenden Morphologie der Speicheldrusenchromosomen // Zool. Jb. 1936. Bd. 56. S. 239-276.
- Zacharias H. Allocyclic behaviour and underreplication of the nucleolus in Pseudodiamesa (Chironomidae). Chromosoma. 1984. 89: 263-273.
- Жимулев И.Ф. Политенные хромосомы: морфология и структура. Новосибирск: ВО Наука. 1992 — 334 с.
- Иванченко О. В. Обзор кариотипов хирономид подсемейств Diamesinae и Prodiamesinae (Diptera, Chironomidae) Палеарктики II. Место и роль двукрылых насекомых в экосистемах. СПб. 1997. С. 54-55.
- Михайлова П. Микроэволюционная дифференциация Pseudodiamesa gr. branickii. /Петрова Н./ Цитология. 1989. Т.31. № 7. С. 824-827.
- Петрова Н. А. Кариотип и неустойчивые ассоциации политенных хромосом Syndiamesa nivosa хирономид (Diptera, Chironomidae). Зоол. журн. 1983. Т. 62. № 1. С. 69-74.
- Петрова Н. А. Характеристика кариотипов хирономид (Diptera, Chironomidae) мировой фауны. I Подсемейства Тelmatogetoninae, Podonominae, Tanypodinae, Diamesinae, Prodiamesinae, Prodiamesinae, Orthocladiinae. Энтомологическое обозрение. 1989. Т. 68. № 1. С. 107-120.
- Петрова Н. А., Кариотипическая характеристика трех примитивных подсемейств хирономид (Diptera, Chironomidae: Tanypodinae, Diamesinae, Prodiamesinae) мировой фауны. / Жиров С. В./ Энтомологическое обозрение. 2013. Т. 92. №3. С.505-516.
- Чубарева Л. А. Кариотипы трех видов таниподин (Tanypodinae, Chironomidae). Тр. ЗИН АН СССР. 1980. Т. 95. С. 65-67[schema type=»book» name=»О НЕОБХОДИМОСТИ РЕВИЗИИ ДИПЛОИДНЫХ ЧИСЕЛ ХРОМОСОМ У НЕКОТОРЫХ ВИДОВ ХИРОНОМИД (DIPTERA, CHIRONOMIDAE)» description=»Пересмотрены диплоидные наборы у трех видов хромосом из рода Pseudodiamesa Goetgh. – Рseudodiamesa stackelbergi Goetgh. (= Sindiamesa nivosa) (2n=8) , Pseudodiamesa ex gr.branickii Nowicki (2n=8) , Pseudodiamesa sp. (2n=8) и вида из рода Anatopinia – A. plumipes Fries (2n=6) на основе данных, полученных при анализе гигантских, митотических и мейотических хромосом. Политенные хромосомы этих видов образуют легко разрушаемый псевдохромоцентр, а отдельно лежащие плечи хромосом образуют различные ассоциации друг с другом, что затрудняет определение хромосомных чисел. На основе статистического анализа частот подобных ассоциаций и данных о недорепликации хроматина в процессе политенизации диплоидные наборы хромосом у указанных видов пересмотрены в сторону уменьшения. » author=»Петрова Нинель Алексеевна, Джапаридзе Людмила Александровна Жиров Сергей Витальевич» publisher=»БАСАРАНОВИЧ ЕКАТЕРИНА» pubdate=»2017-01-19″ edition=»ЕВРАЗИЙСКИЙ СОЮЗ УЧЕНЫХ_28.11.15_11(20)» ebook=»yes» ]