
Бактерия Agrobacteriumtumefaciensнаходит широкое применение в генной инженерии растений благодаря способности переносить свою ДНК в клетки пораженных растений, встраивать ее в геном растения-хозяина, индуцируя стабильную трансформацию клеток собственными генами [1, с. 208-211]. Вирулентные штаммы агробактерий несут Ti-плазмиды, размер которых превышает 200 т.п.о. [2, с. 109-127]. При заражении данным патогеном область Ti-плазмиды длиной 12-25 т.п.н. (Т-ДНК) случайным образом интегрируется в геном пораженного растения, а встроенные гены начинают экспрессироваться. Т-область содержит шесть генов, кодирующих ферментные системы биосинтеза фитогормонов ауксина и цитокинина, их экспрессия приводит к пролиферации клеток и росту опухолевых образований (корончатых галлов) [3, с. 154].
Однако трансформация клеток непосредственно с помощью Ti-плазмиды приводит к образованию опухолевых клеток, из которых невозможно получить целое растение. Поэтому для введения генов используют векторные молекулы на основе Ti-плазмиды с удаленными онкогенами. Вместо онкогенов в вектор по сайтам рестрикции клонируют последовательности целевых генов [4, с. 59-65; 5, с. 289-296].Так, бинарные векторы используют для переноса генетических конструкций, содержащих целевые геныв клетки растений, поскольку в их состав входят соответствующие регуляторные элементы, позволяющие проводить клонирование в E. coli и трансформировать растения с помощью A. tumefaciens. Так, бинарный вектор pBin19 содержит Т-область, фланкированную левой и правой границей (LB и RB), которая в процессе трансформации переносится в геном растения. В Т-ДНК данного вектора расположен также селективный ген nptII, кодирующий последовательность неомицинфосфотрансферазы II и придающий трансформированным клеткам устойчивость к антибиотику канамицину. NptII находится под контролем промотора и терминатора гена нопалинсинтазы (nos), позволяющих экспрессировать данный ген в эукариотических клетках. Для клонирования целевых генов в векторе pBin19 имеется полилинкер, расположенный внутри гена lacZ. Но около полилинкера отсутствует промотор эукариотического типа, поэтому данный вектор не является экспрессирующим для растений. Чтобы клонированные в данном векторе гены могли экспрессироваться, требуется ввести дополнительный промотор, способный функционировать в составе растительного генома.
МАТЕРИАЛЫ И МЕТОДЫ
Для создания модифицированного экспрессирующего вектора использовалась плазмидная ДНКpBin19. Данная плазмида содержит ген nptII, придающий устойчивость к канамицину и находящийся под контролем промотора гена нопалинсинтазы (Pnos), а также полилинкер с множественными сайтами клонирования (рис 1).
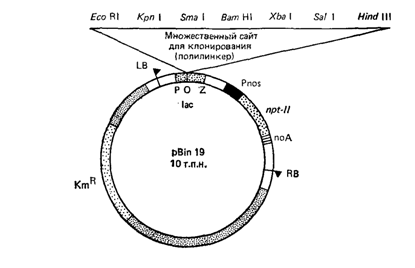
Рисунок 1. Структура бинарного вектора pBin19.
LB, RB – левая и правая границы, lac – α-комплементарная область lac оперона, Pnos – промотор гена нопалинсинтазы, nptII – кодирующая последовательность неомицинфосфотрансферазы II из Tn5, noA – сайт полиаденилирования гена нопалинсинтазы; точками обозначены последовательности плазмиды pRK252, содержащие начало репикации из плазмиды RK2 с широким кругом хозяев.
Для модификации данного вектора использовали нуклеотидные последовательности плазмиды pFF19 (рис. 2).
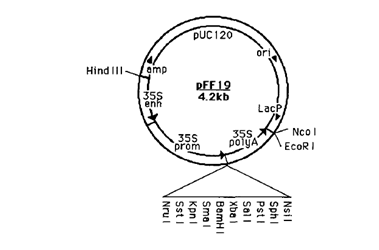
Рис. 2. Карта плазмиды pFF19.
(35Senh – энхансер, 35Sprom — 35S-промотор вируса мозаики цветной капусты, 35SpolyA – сайт полиаденилирования)
Последовательность гена 35S-промотора длиной 994 bp была выделена из плазмидной ДНК pFF19 с помощью эндонуклеаз HindIII и EcoRI. Затем проводили лигирование Т4-лигазой промотора и вектора pBin19 (молярное соотношение вектор/вставка = 1:3) по сайтам рестрикции HindIII и EcoRI с использованием стандартных методик, предложенных фирмой-производителем (СибЭнзим, Новосибирск).
Генетические конструкции на основе вектора pBin19 вводились в клетки E. coli штамма JM 109 методом электропорации с помощью прибора производства Biorad (Bio-Rad Laboratories, США).
Рестрикционный анализ полученных клонов также проводился по стандартным методикам. Результаты анализов визуализировали с помощью электрофореза и обрабатывали с использованием системы гель-документирования GelDoc и программного обеспечения Bio-Rad (Bio-Rad Laboratories, США). В качестве калибровочных маркеров использовали 100 bp +1.5 Kb DNA Ladder (СибЭнзим, Новосибирск).
РЕЗУЛЬТАТЫ И ИХ ОБСУЖДЕНИЕ
Одним из основных регуляторных элементов бинарных векторов является промотор [6, с. 142]. «Сила» промотора, а, соответственно, возможность его использования в генной инженерии, определяется легкостью связывания с промотором фермента РНК-полимеразы и образования открытого комплекса, необходимого для инициации транскрипции [1, с. 108]. Одним из наиболее широко используемых является конститутивный 35S-промотор вируса мозаики цветной капусты (CaMV) [7, с. 285], который активен во всех тканях растения на протяжении всего цикла развития. Так, количественные исследования уровней транскрипции в трансгенных растениях табака показали, что 35S-промотор CaMV, по меньшей мере, в 30 раз сильнее NOS-промотора нопалинсинтазы [8, с. 184].
Последовательность гена 35S-промотора длиной 994 bp была встроена нуклеотидную послеловательность вектора по сайтам рестрикции HindIII и EcoRI (рис. 4 А, Б). На рис. 4 (Б) представлена полученная в результате лигирования конструкция Т-области модифицированного вектора (pBin19mod35S).
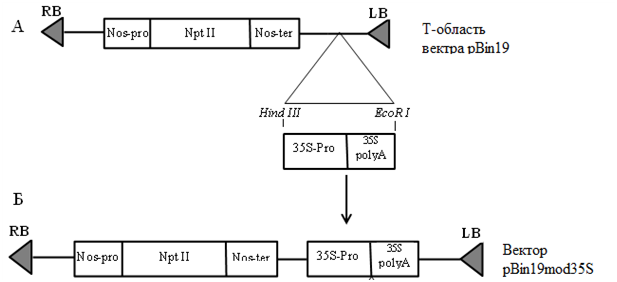
Рис. 3. Схема получения модифицированного бинарного вектора pBin19mod35S
LB, RB – левая и правая границы Т-области, Nos—pro– промотор гена нопалинсинтазы, NptII – кодирующая последовательность неомицинфосфотрансферазы II из Tn5, Nos—ter – сайт полиаденилирования гена нопалинсинтазы; 35S—pro– промотор CaMV, 35S—polyA– сайт полиаденилирования CaMV;
Полученным вектором pBin19mod35S трансформировали клетки E. coli штамма JM109. Рестрикционный анализ полученных клонов на наличие вставки 35S-промотора вируса мозаики цветной капусты проводился по сайтам рестрикции HindIII и EcoRI (рис.4).
Из данной электрофореграммы следует, что в одном из клонов вставка промотора прошла успешно, что подтверждается наличием на треке №6 полосы длиной 994 bp, соответствующей длине последовательности 35S-промотора CaMV. Таким образом, нами был получен экспрессирующий бинарный вектор pBin19mod35S, который можно использовать для клонирования целевых генов, их интеграции в геном растений и эффективной экспрессии в растительных клетках.
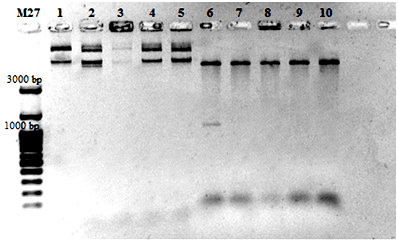
Рис. 4. Электрофореграмма продуктов рестрикционного анализа клонов, полученных в результате трансформации.
Треки 1-5 – нерестрицированная плазмидная ДНК клонов, 6-10 – плазмидная ДНК клонов, обработанная рестриктазами HindIII и EcoRI.
Полученный бинарный вектор был успешно использован для создания генетических конструкций с фрагментами генов транспортного белка и белка оболочки вируса кустистой карликовости малины. Так, фрагменты длиной 503 и 383 bp, соответственно, были клонированы по сайтам рестрикции BamHI и SalIвектора (рис. 5).
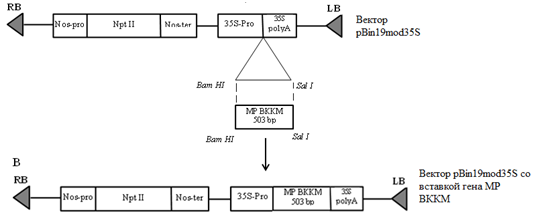
Рис. 5. Схема получения генетической конструкции с фрагментом гена транспортного белка ВККМ
На рисунке 5 представлен вариант получения генетической конструкции на основе фрагмента гена транспортного белка ВККМ. Нами было получено несколько вариантов генетических конструкций. Так, часть конструкций была получена в результате лигирования по одному сайту рестрикции (BamHI илиSalI). Для этого вектор был обработан только одним из ферментов. В результате встраивалось несколько копий целевого фрагмента, что подтверждается результатами рестрикционного и ПЦР-анализа (рис. 6). Из данных рисунка видно, что в результате ПЦР были синтезированы как фрагменты длиной около 503 bp, соответствующие длине целевого фрагмента гена транспортного белка ВККМ, так и фрагменты превышающие длину фрагмента в 4 раза. Также были получены конструкции. Несущие единичные копии целевой последовательности, клонированной по двум сайтам рестрикции.
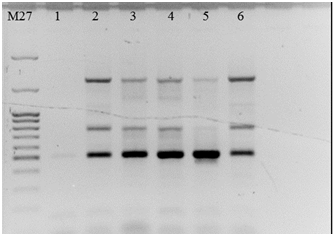
Рис. 6. Электрофореграмма продуктов ПЦР-анализа клонов, несущих фрагмент гена транспортного белка ВККМ
Полученный модифицированный бинарный вектор и созданные на его основе генетические конструкции планируется использовать в дальнейшей работе по получению трансгенных растений малины, устойчивых к вирусу кустистой карликовости малины.
СПИСОК ИСПОЛЬЗОВАННОЙ ЛИТЕРАТУРЫ:
- Патрушев Л. И. Искусственные генетические системы. Т.1. Генная и белковая инженерия. М.: Наука, 2004. – 530 с.
- ZaenenI., VanLarebekeN., VanMontaguM., SchellJ. SupercoiledcircularDNAincrown-gallinducingAgrobacteriumstrains.JMolBiol.,V. 86, № 1, 1974. – P. 109-127.
- Лутова Л.А. Биотехнология высших растений. Санкт-Петербург: СПбГУ,2003. – 243 стр.
- Wahlfors J., loimas S., Pasanen T., Hakkarainen T. Green fluorescent protein (GFP)fusion constructs in gene therapy research.Histochem Cell Biol., Vol. 115. N 1. – P. 59-65.
- Verkhusha V. V., Lukyanov K. A. The molecular properties and applications of Anthozoa fluorescent proteins and chromoproteins. Nat Biotechnol., 22. N3, 2004. – P. 289-296.
- Carter B. T., Lin h., Cornish V. W. Yeast n-hybrid system for molecular evolution. Directed evolution of proteins, or how to improve enzymes for biocatalysis. N.Y. Wiley., – P. 127-158.
- Frank A., Guilley H., Jonard G. Nucleotide sequence of cauliflower mosaic virus DNA. Cell. V. 21., – P. 285-294.
- Harpster, M.H., Townsend, J.A., Jones, J.D.G., Bedbrook, J. Dunsmuir, P. Relative strengths of the 35S cauliflower mosaic virus, 1′, 2′ and nopaline synthase promoters in transformed tobacco, sugarbeet and oilseed rape callus tissue. // Mol. Genet. Vol, 212.,1988. – P. 182-190.[schema type=»book» name=»СОЗДАНИЕ МОДИФИЦИРОВАННОГО БИНАРНОГО ВЕКТОРА ДЛЯ АГРОБАКТЕРИАЛЬНОЙ ТРАНСФОРМАЦИИ РЕМОНТАНТНОЙ МАЛИНЫ» description=»Одним из наиболее эффективных способов борьбы с вирусными инфекциями малины является получение устойчивых трансгенных растений методом агробактериальной трансформации. Для этого целевые нуклеотидные последовательности вводят в клетки A. tumefaciens с помощью бинарных векторов, способных экспрессироваться в прокариотических и эукариотических клетках. Такой бинарный вектор был получен на основе плазмиды pBin19. » author=»Мытницкая Юлия Федоровна, Заякин Владимир Васильевич, Нам Ирина Ян-Гуковна» publisher=»БАСАРАНОВИЧ ЕКАТЕРИНА» pubdate=»2017-03-01″ edition=»ЕВРАЗИЙСКИЙ СОЮЗ УЧЕНЫХ_27.06.2015_06(15)» ebook=»yes» ]