

Дендримеры [1-4] и, в частности, лизиновые дендримеры [5-8], состоящие из натуральных аминокислотных остатков, широко используется во многих промышленных и биомедицинских приложениях. Для улучшения их физико-химических и медико-биологических свойств часто используется модификация их концевых групп другими молекулами. Поэтому необходимо исследование структурных свойств таких гибридов, включая изучение влияния их компонентов на размеры и форму друг друга и системы в целом, а также изучение пространственного расположения этих компонентов в гибриде.
В данной статье нами было проведено моделирование методом молекулярной динамики [9] структуры гибридных молекул, состоящих из лизинового дендримера и дипептидов вилон или тетрапептидов эпиталон, химически пришитых к его концевым группам. Структура дендримера в таких гибридах сопоставлялась со структурой исходного дендримера в воде, а структуры пришитых пептидов в двух гибридах — между собой.
Модель и метод расчета
Методом молекулярной динамики была рассмотрены три системы (Рис.1), состоящие из: 1) из лизинового дендримера третьего поколения. 2) гибрида этого дендримера с 16 дипептидами вилон (LysGlu) , пришитыми к концам дендримера и 3) гибрида этого же дендримера с 16 пептидами эпиталон (AlaGluAspGly), пришитыми к концам дендримера в водном окружении.


Рис.1 Поверхность доступная для внешних контактов в лизиновом дендримере третьего поколения (а), в вилоновом гибриде (б) и в эпиталоновом гибриде (в).
Нескомпенсированный заряд дендримера и «вилонового» гибрида равен +16. Для компенсации заряда этих групп добавляются анионы Cl—. В «эпиталоновом» гибриде общий заряд концевых пептидов дендримера равен -16, поэтому для их компенсации добавляются катионы натрия Na+. В работе использовалась полноатомная молекулярная модель дендримера, пептидов, воды и контрионов и силовое поле AMBER99 и пакет программ для молекулярного моделирования GROMACS. Для уменьшения влияния границ система помещалась в кубическую ячейку с периодическими граничными условиями.
Все системы моделировались в течение 200 нс. Первые 100 нс использовались для установления равновесия, которое контролировалось по установлению постоянной величины радиуса инерции дендримера. Последующие 100 нс использовались для вычисления равновесных характеристик системы (размеров, расстояний и функций распределения).
Результаты
На рис. 1 приведены конформации дендримера и двух гибридов в конце расчета длительностью 200нс. Легко заметить, что размеры всех трех молекул близки, несмотря на значительное различие в их массах. Также хорошо видно, что плотность (заполнение объема) внутренней части эпиталонового гибрида больше, чем у вилонового гибрида, а плотность внутри вилонового гибрида в свою очередь больше, чем плотность аналогичной области исходного дендримера.
Табл. 1 Равновесные значения радиуса инерции Rg, нм.
| Система / подсистема | Rg дендримера (нм) | Rg гибрида (нм) |
| Дендример | 1.14 ± 0.07 | — |
| Гибрид1=Дендример+16 вилонов | 0.99 ± 0.05 | 1.33 ± 0.06 |
| Гибрид2=Дендример+16эпиталонов | 0.93 ± 0.03 | 1.33 ± 0.05 |
Приведенные в таблице 1 среднеквадратичные значения радиусов инерции Rg молекул свидетельствуют о том, химическая пришивка пептидов приводит к увеличению (с 1.14 нм до 1.33 нм) размеров (гибридных систем (т.е. общего радиуса инерции гибрида, состоящего из дендримера и пептидов).
В то же время ковалентная пришивка пептидов к концам дендримера приводит к некоторому уменьшению размеров (радиуса инерции Rg) самого дендримера без учета пришитых к нему пептидов (с 1.14 нм до 0.99 нм для «вилонового» и до 0.93 для «эпиталонового» гибридов). Таким образом несмотря на увеличение общих размеров гибридов на 17%, их внутренняя часть поджимается по сравнению со свободным дендримером на 15% и 23% для вилонового и эпиталонового гибридов соответственно.
Функция распределение дендримеров и гибридных молекул по радиусам инерции, а также соответствующие распределения для главных компонент тензора инерции позволяют судить об амплитуде флуктуаций этих величин и анизотропии формы дендримера и гибридов. На рис.2 приведены фунции распределения для этих величин в вилоновом (рис.2а) и в эпиталоновом (рис.2б) гибридах. Сравнение распределения Rg гибридов показывает, что не только средние значения Rg, но и функции распределения по радиусам инерции гибридов близки (обе флуктуируют в основном между значениями 1.2 и 1.5 нм), несмотря на то, что тетрапептид эпиталон примерно вдвое тяжелее, чем дипептид вилон.
Функции распределения для главных компонент Rg отличаются друг от друга меньше для эпиталонового гибрида, что говорит о близкой к сферической форме этого гибрида. Для вилонового гибрида распределение для одной (меньшей) компоненты сдвинуто в сторону меньших величин Rg и больше (хотя и не слишком сильно) отличается от двух других компонент Rg, чем в эпиталоновом гибриде. Это говорит о том, что вилоновый гибрид имеет форму промежуточную между дискоообразной и сферической.
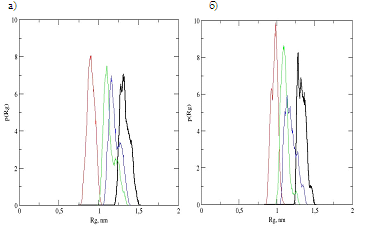

Рис. 2. Функции распределения вилонового (а) и эпиталонового (б) гибридов по радиусам инерции. Черные линиии – для радиуса инерции, цветные – для главных компонент тензора инерции.
Более детальную информацию о распределении атомов в гибридах дают функции распределения атомов по расстояниям от центра инерции дендримера (см. рис.3).
На рис.3а сопоставлены соответствующие функции для дендримера (черная линия) и дендримерной части вилонового (красная линия) и эпиталонового (зеленая линия) гибридов.
Легко видеть что максимум распределения атомов в дендримерной часть обоих гибридов смещается к меньшим значениям расстояния до центра инерции по сравнению с исходным дендримером. При этом пришивка более массивных и обемных тетрапептидов эпиталон (зеленая линия) поджимает дендример сильнее, чем пришивка менее массивных и объемных дипептидов вилон. Таким образом, пришивка более массивных пептидов приводит к большему увеличению плотности внутренней дендримерной части гибрида. Эти данные хорошо коррелируют с данными об уменьшении радиусов инерции Rg дендримерной части гибридов, приведенными в табл.1.
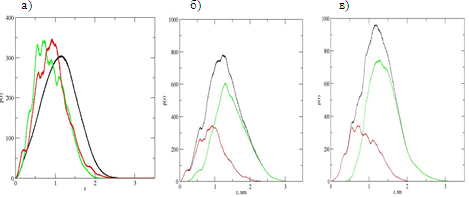

Рис.3а) радиальная функция распределения p(r) атомов одной и той же дендримерной части (без учета атомов пептидов) относительно центра инерции в разных системах: а) в дендримере (черная линия), вилоновом (красная линия)) и эпиталоновом (зеленая линия) гибридах.
Рис 3б и 3в: Функция распределения p(r) различных групп атомов в каждом из двух гибридов: б) в вилоновом» гибриде и в) в эпиталоновом гибриде». Все атомы гибрида (черные линии), атомы самого дендримера в гибриде (красные линии) и атомы пептидов в гибриде (зеленые линии) в зависимости от расстояния до центра дендримера r.
На рис. 3б и рис. 3в построены соответствующие функции распределения для всех атомов каждого из гибридов и отдельно для атомов входящих в гибрид дендримера и пептидов. Пик функции распределения всех атомов в гибридных молекулах с пришитыми пептидами смещается на большие расстояния по сравнению с исходным дендримером, что коррелирует с увеличением средних размеров гибрида по сравнению с дендримером (таблица 1). Из сравнения распределения атомов пришитых пептидов относительно центра дендримера в каждом из гибридов с распределением атомов самого дендримера в соответствующем гибриде (рис.3) можно сделать вывод, что пептиды находятся в основном на поверхности дендримера или недалеко от нее и, обычно, не проникают глубоко внутрь дендримера. В то же время сравнение проникновения вилонов на рис.3б и эпиталонов на рис.3в к центру соответстующих гибридов показывает, что более легкие вилоны проникают несколько глубже, чем более массивные и объемные эпиталоны. Этот результат находится в согласии с ранее установленными (в том числе и в работах авторов) общими закономерностями для дендримеров с объемными концевыми группами, для которых проникновение концов внутрь дендримера (backfolding) затруднено.
Важной характеристикой заряженных дендримеров (и гибридов) является распределение заряженных групп. В исходном лизиновом дендримере третьего поколения имеется 16 положительно заряженных концевых групп NH3+. В вилоновом гибриде имеются 32 NH3+ на концевых лизинлвых аминокислотных остатках вилона и 16 отрицательно заряженных COO— групп на предшествующих остатках глутаминовой кислоты, так что общий заряд концов дендримера остается тем же (+16), что и в исходном дендримере, а эпиталоновом гибриде имеется 16 концевых NH3+ групп и 32 отрицательно заряженные COO— группы, расположенные влизи от концов, так что суммарный заряд на трех терминальных аминокислотных остатках эпиталона в гибриде равен -16. Мы рассчитали из молекулярно-динамического моделирования среднюю фукцию распределения положительных и отрицательных зарядов в обоих гибридах, а также распределение общего заряда в зависимости от расстояния до центра дендримера (см. рис.4а и рис.4б).
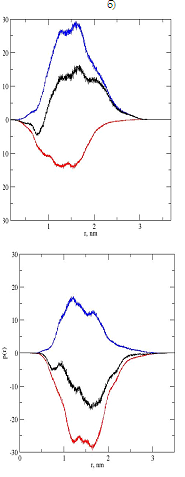

Из рис.4а и 4б видно, что распределение положительного и отрицательного зарядов смещены относительно друг друга. В случае вилонового гибрида (рис.4а) это приводит к небольшому преобладанию отрицательных зарядов на малых расстояниях от центра (0.7-0.9 нм), приблизительно нулевому заряду на расстоянии порядка 1 нм от центра и существенному преобладанию положительного заряда на расстояниях от 1 до 2.5 нм.
Для эпиталонового гибрида отрицательный заряд преобладает на всех расстояниях начиная от 0.5 нм и до 2.5 нм.
Для оценки поступательной подвижности дендримера и гибридов в воде были построены временные зависимости среднеквадратичных смещений центров инерции дендримера и гибридов в воде (см. рис.5).
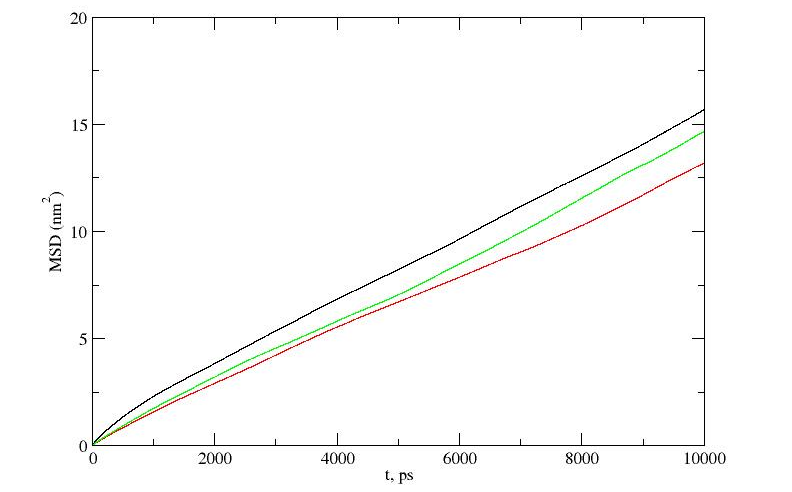

Зависимости этих смещений от времени оказались во всех случаях близкими к линейным, что свидетельствует о достижении диффузионного режима движения. Из наклона этих зависимостей были определены коэффициенты поступательной диффузии дендримера и гибридных молекул (см. Табл.2).
Табл. 2. Коэффициенты поступательной диффузии D дендримера и гибридов.
| Система | D (см2/с) x105 |
| Дендример | 0.26 ±0.03 |
| Гибрид1=Дендример+16 вилонов | 0.24 ± 0.02 |
| Гибрид2=Дендример+16 эпиталонов | 0.22 ± 0.08 |
Было получено, что коэффициент поступательной диффузии вилонового гибрида в воде меньше, чем у эпиталонового гибрида, который в свою очередь меньше коэффициента диффузии исходного лизинового дендримера третьего поколения. Однако эти различия не слишком велики и составляют примерно 10% при переходе от одной молекулы к другой. Таким образом подвижность всех трех молекул в воде оказывается довольно близкой, несмотря на довольно существенные различия в их массах. Это означает, что гидродинамические радиусы рассмотренных гибридов, которые можно оценить из полученных коэффициентов диффузии, отличаются от гидродинамического радиуса исходного дендримера гораздо меньше, чем это можно было ожидать исходя из суммы среднеквадратичных размеров компонент гибридов (дендримера и пептидов). Эти данные коррелируют с полученным нами ранее относительно небольшим увеличением радиусов инерции Rg гибридов по сравнению c Rg исходного дендримера.
Выводы:
- Методом молекулярной динамики промоделированы гибридные молекулы, состоящие из лизинового дендримера третьего поколения и пришитых к нему 16 дипептидов вилон или 16 тетрапептидов эпиталон.
- Показано, что пришивка пептидов к дендримеру приводит к ожидаемому увеличению размера совокупной системы. В то же время оказывается, что размеры самого дендримера при этом несколько уменьшаются.
- Пришитые пептиды находятся в основном на поверхности дендримера или вблизи нее и таким образом доступны для взаимодействий с другими молекулами как в растворе, так и в организме в случае использования гибридной структуры в качестве средства доставки лекарственных препаратов. При этом для вилонового гибрида это свойство менее выражено, чем для эпиталонового.
- Распределение заряженных групп существенно отличается в вилоновом и эпиталоновом гибриде. В случае вилонового гибрида общий заряд дендримера (+16) и концевой аминокислотный остаток (Lys) не меняется по сравнению с исходным лизиновым дендримером, что приводит к распределению суммарного заряда сходному с распределением в исходном лизиновом дендримере. В то же время в эпиталоновом гибриде, несмотря на наличие 16 положительно заряженных концевых групп, общий заряд (-16) отрицателен. Это должно приводить к принципиальному отличию биомедицинских свойств и области возможного применения последнего гибрида. В частности можно предположить, что эпиталоновые гибриды в отличие от лизиновых дендримеров и вилоновых гибридов должны гораздо хуже проникать сквозь липидные бислои.
- Коэффициент поступательной диффузии вилонового гибрида в воде оказывается меньше, чем у эпиталонового гибрида, который в свою очередь меньше коэффициента диффузии исходного лизинового дендримера третьего поколения. Однако, эти различия невелики (<10%) , что означает небольшое отличие гидродинамических радиусов рассмотренных систем, которое хорошо коррелируют с небольшим увеличением радиусов инерции Rg гибридов по сравнению с Rg исходного дендримера.
Эта работа была частично поддержана грантами РФФИ 16-03-00775 и 15-33-20693
mol_a_ved и грантом Правительства Российской Федерации, 074-U01.
Список литературы:
- Klein A.V., Fréchet J. M. J., Tomalia D. A. Dendrimers and other dendritic polymers, Chichester: Eds. Wiley, 2001.- 513p.
- Dendrimers in Biomedical Applications, ed. B. Klajnert, L. Peng and V. Cena, RSC Publishing, 2013.
- Mazo M. A., Shamaev M. Yu., Balabaev N. K., Darinskii A. A., Neelov I. M. // Phys.Chem.Chem.Phys., 2004, 17, 1285–1289.
- Neelov I.M. , Adolf D.A. Macromolecules 2003, 36, 6914-6924
- Falkovich S., Markelov D., Neelov I. and A. Darinskii // J. Chem. Phys., 2013, 139, 064903.
- Neelov I. M., Markelov D. A., Falkovich S. G., Ilyash M. Y., Okrugin B. M. and Darinskii A. A.// Polym. Sci., Ser. C, 2013, 55, 154–161.
- Neelov I.M., Janaszewska A., Klajnert B., Bryszewska M., Makova N.Z., Hicks D.A., Pearson H.A., Vlasov G.P., Ilyash M.Y., Vasilev D.S., Dubrovskaya N.M., Tumanova N.L., Zhuravin I.A., Turner A.J., Nalivaeva N.N.// Current Medicinal Chemistry, 2013, Vol. 20, No. 1, pp. 134-143.
- Markelov D., Falkovich S., Neelov I., Ilyash M., Matveev V., Lahderanta E., Ingman P. and Darinskii A. // Phys.Chem.Chem.Phys., 2015, 17, 3214–3226.
- Gotlib Yu.Ya., Balabaev N. K., Darinskii A. A., Neelov I. M. // Macromolecules 1980, 13, 602-608.[schema type=»book» name=»Молекулярно-динамическое моделирование лизиновых дендримеров и их гибридов с биоактивными пептидами » description=»Целью данной работы было установление структурных характеристик лизинового дендримера и его гибридов с короткими биоактивными пептидами в разбавленном водном растворе методом молекулярной динамики. Было получено, что химическая пришивка биоактивных пептидов в гибридах ко всем терминальным группам лизинового дендримера приводит к небольшому увеличению размеров системы и увеличению ее внутренней плотности. При этом сам дендример оказывается поджатым, а пришитые пептиды располагаются в основном на поверхности дендримера и лишь немного проникают внутрь него.» author=»Ильяш Максим Юрьевич, Округин Борис Михайлович, Хамидова Дилором Насруллоевна, Неелов Игорь Михайлович» publisher=»БАСАРАНОВИЧ ЕКАТЕРИНА» pubdate=»2016-12-21″ edition=»euroasian-science.ru_25-26.03.2016_3(24)» ebook=»yes» ]