

Пустынно — степные условия юга Казахстана отличаются особо сложным ко-мплексом неблагоприятных факторов, влияющих на растительные организмы: интенсивная засушливость климата, необычная жара летом, крайне низкая от-носительная влажность воздуха, поздние весенние заморозки, незначительный снежный покров, разной степени засоление почвы и глубокое залегание грунто-вых вод, часто минерализованных. Все это создает трудности в осуществлении интродукции в эти районы. Интродукция растений в значительной степени яв-ляется эколого-географической проблемой [4,-С.78].
Способность растений изменяться в новых условиях их произрастания яв-ляется предпосылкой возникновения новых адаптивных качеств. Это зависит от особенностей растительных организмов, сложившихся на протяжении длитель-ного эволюционного их развития [6,-С. 8].
Приспособление древесных растений к перенесению засухи идет у разных видов различными путями.
Экология интродуцированных растений аридных зон юга Казахстана изу-чена крайне недостаточно. Отсутствуют сведения об анатомических особен-ностях и степени приспособленности интродуцентов, а также степени их эко-логической изменчивости и пластичности. Знание структурных особенностей этих растений позволит выявить сложные пути их адаптации к жестким усло-виям окружающей среды.
Материалы проведенного исследования обеспечат научно-обоснованный по-дход в прогнозировании выбора древесных и кустарниковых пород для интро-дукции и позволит расширить озеленительный ассортимент к аридным усло-виям предгорий Каратау.
По методике Прозиной М.Н. [7,-С.19-43] нами были исследованы анатоми-ческое и морфологическое строение листьев гледичии трехколючковой и ивы белой. Так как лист более пластичен, чем другие органы растения, и, следо-вательно аридность его местообитания прежде всего сказывается на его стро-ении [1,-С.201-207].
Лист легко изменяет свою структуру не только под непосредственным вли-янием среды, но и при воздействии происходящих внутри растения процессов, возникающих также под влиянием внешних условий. Лист более чутко, чем другие органы растения, перестраивает свою морфолого – анатомическую стру-ктуру, приспосабливаясь к изменению внешней среды.
При уменьшении увлажнения структура листа изменяется в сторону увели-чения более ксероморфных признаков, что сказывается не только в сокращении площади листа, но и в увеличении длины сети жилок и количества устьиц на единицу его площади [2,-С. 75-86].
Лист гледичии трехколючковой – Gleditschia triacanthos L. изолатераль-ный, гипостоматический. Мезофилл листа состоит из очень мелких, прибли-зительно одинакового размера клеток.
Палисадная паренхима, расположенная под верхним эпидермисом состоит из 6 слоев плотносомкнутых клеток клеток. Толщина его 55±1-2 мкм.
Среднюю часть мезофилла занимает губчатая паренхима, состоящая из 4 сло-ев клеток. Клетки губчатой паренхимы расположены довольно плотно. Меж-клетники развиты слабо. Нижняя палисада состоит из 3-4 слоев мелких клеток, но они расположены не так плотно, как клетки верхней палисады. Процент па-лисадности от толщины мезофилла составляет 61,8%.
Клетки верхнего эпидермиса сравнительно крупные, отличаются толщиной своих стенок и составляют 8,2 мкм. Они имеют форму 3-5 –угольных неправи-льных многоугольников со слегка извилистыми стенками. Устьица отсутст-вуют. Имеются простые волоски. Длина волосков от 6,96 до 17,4 мкм.
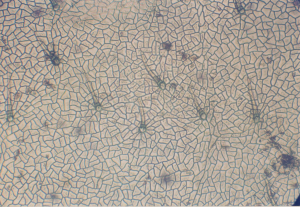

Рисунок 1. Верхний эпидермис листа гледичии трехколючковой
Клетки нижнего эпидермиса намного мельче. Их форма не меняется, но ко-личество соответственно значительно увеличивается до 4700 на 1 мм2 устьица мелкие, но расположены друг от друга нечасто. На нижней стороне листовой пластинки волоски отсутствуют. Стенки клеток слегка извилистые, их толщина 1,5 мкм.
Устьица и главная жилка погружены в толщу листа. Погруженность устьиц и главной жилки в толщу пластинки листа, имеющая явное приспособительное
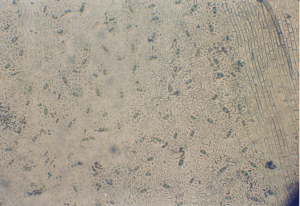

Рисунок 2. Нижний эпидермис листа гледичии трехколючковой
значение, могла выработаться в течение относительно долгих сроков, но, веро-ятно, соизмеримых со сроками существования тех или иных аридных областей.
К этой же категории признаков относятся и изолатеральный мезофилл.
Распространенность этих признаков среди пустынных растений дает осно-вание заключить, что обладание ими дает виду некоторые существенные пре-имущества в борьбе за существование в условиях засушливых местообитаний.
Учитывая анатомические признаки листа гледичии ее можно отнести к ксе-рофитам.
Лист ивы белой – Salix alba L.- изолатеральный, амфистоматический.
Мезофилл состоит из 5 рядов плотносомкнутых клеток палисадной парен-химы и 1 слоя гиподермы. Своеобразной чертой анатомии листа Ивовых явля-ется наличие в нем гиподермы, характерной для многих, хотя не для всех видов этого семейства. Она состоит из бедных содержимым, как бы пустых, лишен-ных хлоропластов клеток с оболочкой обычно несколько более толстой, чем у остальных клеток хлоренхимы. Если не обратить внимание на отсутствие в этих клетках хлорофилла и на довольно резкое отличие в окрашиваемости сте-нок гематоксилином, гиподерму можно принять за часть губчатой паренхимы.
Esau определяет гиподерму как специализированную субэпидермальную тка-нь, которая может быть частью окружающей ее хлоренхимы или возникает из протодермы. У ив гиподерма функционально и вероятно, генетически не впол-не обособлена от хлоренхимы. В нормально сформированных листьях ив в кон-це лета однослойная гиподерма всегда содержит хлоропласты. Слой гиподермы расположен под нижним эпидермисом листовой пластинки.
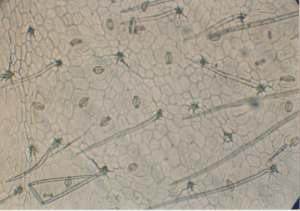

Рисунок 3. Нижний эпидермис листа ивы белой
Клетки мезофилла вытянутые, узкие, их длина в 3-4 раза превышает ширину. Межклетники отсутствуют. Снаружи лист покрыт эпидермой и кутикулой. Ку-тикула эпидермиса многих ивовых образуют своеобразные утолщения в виде продольных валиков, которые на поперечном разрезе листа выглядят как зубцы или гребешки. Подобные скульптурные образования характерны для кутикулы нижнего эпидермиса многих ив. Общая толщина листовой пластинки состав-ляет 348,0±3,52 мкм.
Клетки как верхнего так и нижнего эпидермисов многогранные, таблитчатой формы. Стенки клеток прямые, неизвилистые. Высота клеток эпидермиса 66,3 мкм. Толщина стенок верхнего эпидермиса составляет 5,5 мкм, нижнего 1,5 мкм. Имеются простые, довольно длинные одноклеточные волоски, располо-женные по всей поверхности верхнего эпидермиса. Количество волосков в поле зрения микроскопа 25 штук. Эпидермис у ив имеет прямые антиклинальные стенки, что находит выражение в так называемой таблитчатости его клеток. Особенно заметна таблитчатость при рассмотрении листа с поверхности. Эта особенность строения свойственна клеткам и верхнего и нижнего эпидермиса ив. На нижнем эпидермисе волоски отсутствуют.
Устьица располагаются на обеих сторонах листовой пластинки. На верхнем эпидермисе их меньше, чем на нижнем. Устьица аномоцитного типа.
Данную структуру листа вполне можно назвать ксероморфной.
Скворцов А.К., Голышева М.Д. [7,-С.91-97], изучая особенности строения листьев рода Salix L., и сопоставив полученные результаты с данными Васи-левской В.К., [5,-С.5-17] Александрова В.Г., Мирославова Е.А. [3,-С.852-856] убедились в большой константности признаков анатомической структуры листа рода Salix L., свойственной таксонам различного ранга.
Наши исследования полностью подтверждают эти данные. Особенно харак-терно строение поперечного сечения и строение эпидермиса.
Александров В.Г., и Мирославов Е.А. [3,-С.852-856], изучая особенности структуры листьев некоторых видов ив, произрастающих на северо-западе СССР, утверждают, что «ни одна из ив Ленинградской области не произрастает в условиях, которые бы способствовали развитию ксероморфной структуры листьев; все ивы произрастают не только при достаточном, но даже избыто-чном увлажнении почвы. Листья собраны с растений, произрастающих в условиях, исключающих ксерофитные».
Таким образом, изученный нами вид ивы белой, интродуцированный в пред-горную аридную зону Каратау, обнаруживает совершенно сходное строение ли-ста.
Список литературы
- Александров В.Г., Цхакая К.Е. К проблеме степени пластичности листа и о возникновении ксероморфной структуры. //Тр. с/х. — Опытного учрежд. Дона и Сев. Кавказа.1926. –С.201-207
- Александров В.Г., Александрова О.Г., Тимофеев С.А. Водоснабжение ли-ста и его строение.//Зап. научн. прикл. отд. Тифл. Бот. сада. -1921. вып.2. – С.75-86
- Александров В.Г., Мирославов Е.А. Об особенностях структуры листьев некоторых видов ив, произрастающих на северо–западе СССР. //Бот.журнал.1962.т.XVII.№ –С.852-856
- Байтулин И.О., Проскуряков М.Л., Шкалин С.В. Системно – экологиче-ский подход к интродукции растений в Казахстане. ч.1-2. Алма –Ата. 1992. –С.78
- Василевская В.К. Формирование листа засухоустойчивых растений. Ашхабад. 1954. –С.5-17.
- Коновалов И.Н. Физиология интродуцируемых растений. М-Л., 1963. –С.8-
- Прозина М.Н. Ботаническая микротехника. М.1960. –С.19-43
- Скворцов А.К., Голышева М.Д. О некоторых особенностях строения ли-ста, важных для систематики и филогении рода Salix L. //Биол. науки. 1967. №5.-С.91-97.[schema type=»book» name=»ИНТРОДУКЦИЯ ДРЕВЕСНЫХ РАСТЕНИЙ В АРИДНЫЕ ЗОНЫ ЮГА КАЗАХСТАНА» description=» В данной статье рассматриваются проблемы интродукции и акклиматизации древесных растений в аридные зоны юга Казахстана, изучив морфологическое и анатомическое стро-ение листьев исследуемых растений. » author=»Рысмамбетова Г.М., Байжигитов Д.К., Оразбаева О.О.» publisher=»БАСАРАНОВИЧ ЕКАТЕРИНА» pubdate=»2017-03-30″ edition=»ЕВРАЗИЙСКИЙ СОЮЗ УЧЕНЫХ_30.04.2015_04(13)» ebook=»yes» ]