

Методы:
В работе использованы реактивы фирмы Sigma (США). В качестве экспериментальных животных служили самцы и самки белых беспородных крыс 6-и и 10-и недельного возраста. Раствор цисплатина (Cis-DDP) вводили внутрибрюшинно (10 мг/кг веса животного). Животных декапитировали через 48 часов после инъекции препарата. Выделение ядер производили по модифицированному методу Hewish at al [3, с. 505].
Ядра клеток печени осаждали центрифугированием при 24000 g в течение 60 мин. Осадок очищенных ядер отмывали от избытка сахарозы в среде выделения с последовательным осаждением (центрифугирование 10 мин при 700 g).
Активность ПАРП 1 измеряли по убыли субстрата (НАД+) в реакционной среде, содержащей изолированные ядра клеток печени (плотность ядерной суспензии составляла 1мг/мл по ДНК), 1мМ CaCl2, 6 мМ MgCl2 в течение 10 мин. Реакцию инициировали добавлением НАД+ к суспензии ядер. Через 10 мин реакцию останавливали путем осаждения ядер из реакционной среды центрифугированием (2 мин 8000 g, 370C). Количество остаточного НАД+ определяли в образцах надосадочной жидкости (50 мкл) по методу, основанному на колориметрическом определении окрашенного продукта реакции НАД+ с ацетофеноном (поглощение 378 нм) [4, с. 80]. Количество НАД+ определяли по калибровочной кривой.
Методом вестерн блоттинга идентифицировали ПАРП 1 и фрагменты специфического расщепления фермента. После переноса фракций электрофоретического разделения ядерных белков мембрану инкубировали в блокирующем растворе (10 мМ Трис HCl, pH 7,4; 150 мМ NaCl; 0,05% Tween 20). Для предотвращения неспецифического связывания антител в инкубационный раствор добавляли бычий сывороточный альбумин (BSA) и инкубировали при + 4°C, в течение ночи. Далее мембрану промывали 3 раза по 5 мин в инкубационном растворе (10 мМ Трис HCl, pH 7,4; 150 мМ NaCl; 0,05 % Tween 20).
Первичные антитела ПАРП-1 вносили в раствор для инкубации мембран (1:2500). Через 2 часа (+4°C) несвязавшиеся первичные антитела удаляли промывкой мембраны инкубационным раствором. Далее мембрану выдерживали в течение 1 часа в инкубационном растворе, содержащем вторичные антитела (1:25000), конъюгированные со щелочной фосфатазой.
Достоверность различий между средними величинами полученных данных определяли по t-критерию Стьюдента.
Результаты:
В работе исследовалось действие цисплатина и аллостерического ингибитора поли(АДФ-рибозо)полимеразы 1 (ПАРП 1) АТФ на активность фермента в ядрах клеток печени самцов и самок 6-и и 10-и недельных крыс.
Полученные данные выявили заметные половые различия в активности ПАРП 1. Было показано, что в ядрах клеток печени неполовозрелых крыс самцов (6 нед.) базовая активность ПАРП 1 на 40% выше активности фермента печени крыс самок (рис. 1).
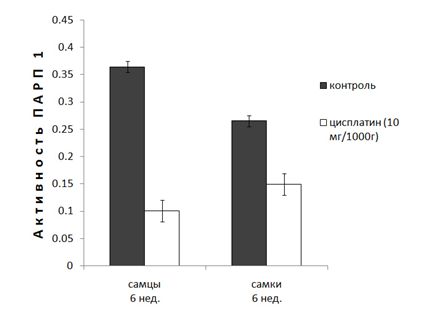

Рисунок 1. Действие цисплатина (10мг/кг, 48ч) на активность ПАРП 1 в ядрах клеток печени неполовозрелых (6 нед.) крыс.
В ядрах клеток печени половозрелых крыс (10 нед.) различия в активности ПАРП 1 у самок и самцов не наблюдаются. Однако у 10-и недельных крыс наблюдается уменьшение базовой активности фермента в 2.5-3 раза (рис. 2). Эксперименты показали, что после введения цисплатина 6-и недельным животным, активность ПАРП 1 самцов и самок уменьшается в различной степени (3,5 и 2 раза, соответственно). Однако введение цисплатина 10-и нед. крысам приводит к увеличению активности ПАРП 1 более чем в 2 раза независимо от пола животных (рис 2), что является ярким проявлением возрастных различий действия цисплатина на активность фермента.
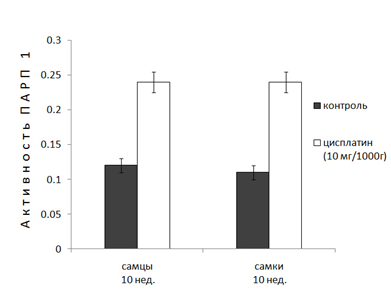

Рисунок 2. Действие цисплатина (10мг/кг, 48ч) на активность ПАРП 1 в ядрах клеток печени половозрелых (10 нед.) крыс.
Во второй серии экспериментов исследовалось влияние различных физиологических концентраций аллостерического ингибитора АТФ на активность ПАРП 1 в ядрах клеток печени. Результаты исследований показали, что у неполовозрелых крыс АТФ (1мМ) ингибирует активность фермента на 40%. У 10-и нед. животных проявляется половой диморфизм действия 1мМ АТФ (активность ПАРП 1 в ядрах печени самок уменьшается на 45%, у самцов-на 10%). Более высокие физиологические концентрации АТФ (5мМ и 10 мМ) практически полностью подавляют активность ПАРП 1 во всех исследуемых группах (рис. 3).
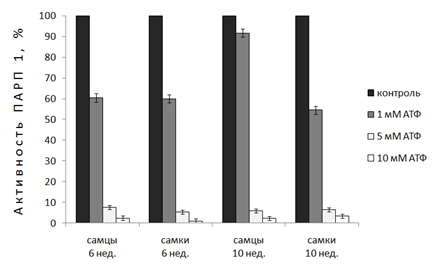

Рисунок 3. Действие физиологических концентраций АТФ на активность ПАРП 1 в ядрах клеток печени неполовозрелых (6 нед.) и половозрелых крыс (10 нед.).
Введение цисплатина 6-и нед. крысам не влияет на степень ингибирования ПАРП 1 АТФ. У 10-и нед. самцов цисплатин увеличивает эффективность ингибирования фермента 1мМ АТФ, нивелируя половые различия в ингибировании (рис. 4).
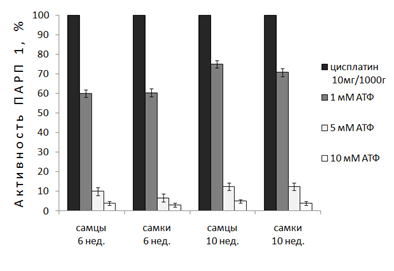

Рисунок 4. Действие физиологических концентраций АТФ на активность ПАРП 1 в ядрах клеток печени неполовозрелых (6 нед.) и половозрелых (10 нед.) крыс после 48ч введения цисплатина (10мг/кг).
Методом вестерн блоттинга было показано, что резкая инактивация ПАРП 1 после добавления АТФ в среду инкубирования ядер клеток печени крыс не обусловлена протеолитическим расщеплением фермента (рис. 5).


Рисунок 5. Идентификация ПАРП 1 в ядерных белках клеток печени крыс. а) ядра инкубировали в среде, не содержащей АТФ б) ядра инкубировали в среде, содержащей 5мМ АТФ
Выводы:
- Активность ПАРП 1 в ядрах клеток печени крыс значительно уменьшается в процессе полового созревания.
- Введение цисплатина крысам исследованных возрастных групп вызывает разнонаправленное изменение активности ПАРП 1. У крыс пубертантного возраста (6 нед.) препарат заметно подавляет активность фермента, в то время как активность ПАРП 1 в ядрах печени 10-и нед. крыс увеличивается в 2,5 раза.
- Высокие физиологические концентрации АТФ (5мМ и 10мМ) полностью подавляют активность ПАРП 1 в ядрах клеток печени крыс. 1мМ АТФ подавляет активность ПАРП 1 в разной степени у 10-и недельных самок и самцов, проявляя половой диморфизм в ингибировании фермента.
- У 10-и недельных самцов цисплатин увеличивает эффективность ингибирования фермента 1мМ АТФ, нивелируя половые различия в ингибировании.
- Резкое подавление активности ПАРП 1 в ядрах клеток печени крыс при воздействии высоких физиологических концентраций АТФ не обусловлено расщеплением фермента.
Список литературы:
- D’Amours D., Desnoyers S., D’Silva I. and Poirier G.G. Poly(ADP-ribosyl)ation reactions in the regulation of nuclear functions. Biochem. J., 1999, 342: 249-268.
- Me´gnin-Chanet , M.A. Bollet ,J. Hall/Targeting poly(ADP-ribose) polymerase activity for cancer therapy/Cell. Mol. Life Sci., 2010, 67: 3649–3662.
- Hewish D.R., Burgoyne L.A. Chromatin Substructure, The Digestion of Chromatin DNA at Regularly Spaced Sites by a Nuclear Deoxyribonuclease. Biochemical and Biophysical Res. Com, 1973, 52: 504-510.
- Putta S. and Hergenrother P.J. An enzymatic assay for poly(ADP — ribose) polymerase – 1 (PARP-1) via the chemical quantitation of NAD+ : application to the high-throughput screening of small molecules as potential inhibitors. Analytical Biochemistry, 2004, 326: 78 – 86.
- Rouleau M., Aubin R., Poirier G. Poly(ADP-ribosyl)ated chromatin domains: access granted. Journal of Cell Science, 2004, 117(6), 815-825.
- Szabo C.., Pacher. P., . Swanson R. A. Novel modulators of poly(ADP-ribose) polymerase Trends Pharmacol Sci. 2006; 27(12): 626–630.
- Zhu G., Chang P., Lippard S.. Recognition of Platinum-DNA Damage by Poly(ADP-Ribose)Polymerase-1, J. Biochemistry , 2010; 49(29), p.6177-6183 .
«This work was made possible by a research grant from the Armenian National Science and Education Fund (ANSEF) based in New York, USA»[schema type=»book» name=»ДЕЙСТВИЕ ЦИСПЛАТИНА НА АКТИВНОСТЬ ПОЛИ(АДФ-РИБОЗО) ПОЛИМЕРАЗЫ 1 В ЯДРАХ КЛЕТОК ПЕЧЕНИ КРЫС» description=»Опухолеобразование является сложным и многоэтапным процессом, чаще всего обусловленным бесконтрольным ростом той или иной клеточной популяции вследствие нарушения молекулярных процессов, контролирующих и регулирующих нормальное протекание клеточной пролиферации. Наряду с этим развитие злокачественных новообразований во многих случаях обусловлено нарушениями и неисправностями клеточных механизмов, ответственных за элиминацию перерожденных клеток. Мишенью многих препаратов, которые в настоящее время широко применяются в лечении злокачественных новообразований, является ядерная ДНК раковых клеток. Одним из наиболее эффективных химиотерапевтических ДНК-алкилирующих препаратов является цисплатин. Эффективность цисплатина связана с повреждением ДНК опухолевой клетки, в результате чего формируются цис-ДДП–ДНК–аддукты, блокирующие репликацию, транскрипцию и клеточную пролиферацию в целом [2, с. 3556]. Поли(АДФ-рибозо)полимераза 1 (ПАРП 1), будучи мощным эпигенетическим регулятором, играет ключевую роль в различных механизмах клеточной гибели [5, с. 822]. В ответ на повреждения ДНК ПАРП 1 активируется более чем в 500 раз [1, с. 250], тем самым способствуя успешной сборке репарационной системы в точках ДНК-разрывов, что в конечном итоге приводит к выживанию клеток [7, с. 6181]. Таким образом, инактивация ПАРП 1 увеличивает цитотоксическое действие цисплатина на опухолевые клетки. Данные клинических испытаний указывают на проявление половых различий в терапевтическом эффекте ингибиторов ПАРП 1 [6, с. 627]. Актуальной задачей современной медицины является исследование взаимодействия лекарств при сочетанном применении в сложных химиотерапевтических схемах лечения опухолей. Целью настоящей работы является сравнительное изучение действия терапевтических доз цисплатина на активность и ингибирование ПАРП 1 аллостерическим ингибитором АТФ в клетках печени крыс разного пола и возраста. » author=»Асатрян Ануш Левоновна » publisher=»БАСАРАНОВИЧ ЕКАТЕРИНА» pubdate=»2017-02-02″ edition=»ЕВРАЗИЙСКИЙ СОЮЗ УЧЕНЫХ_26.09.15_10(18)» ebook=»yes» ]